

Você já notou que algumas plantas de soja conseguem manter produtividade elevada mesmo com densidade reduzida, enquanto outras sucumbem à competição intraplantas? A resposta está em um atributo genético-fisiológico denominado plasticidade fenotípica—a capacidade de alteração morfológica em resposta ao ambiente.
Enquanto você investe em tecnologia de ponta no caule principal, 25-40% da produtividade está sendo alocada em ramos laterais mal desenvolvidos. A genética define o potencial máximo; o ambiente (luz, nutrição, água) define a eficiência real. Uma simples reconfiguração de população de semeadura, combinada com manejo nutricional direcionado e aplicação estratégica de bioativos, pode liberar 2-3 sacas “escondidas” em cada hectare.[1][2]
Este artigo apresentará os fundamentos fisiológicos da ramificação lateral em soja, os fatores ambientais que limitam seu desenvolvimento produtivo, e as estratégias comprovadas de otimização—incluindo aplicação de reguladores de crescimento naturais e sintéticos—que você pode implementar imediatamente. Você descobrirá como a plasticidade da soja permite compensação entre densidade de plantas e arquitetura individual, e aprenderá a explorar este mecanismo através de decisões integradas de genótipo, população, nutrição e bioativos.
Plasticidade Fenotípica da Soja: Conceitos Fundamentais
A plasticidade fenotípica refere-se à capacidade de uma planta alterar sua morfologia e fisiologia em resposta a variações ambientais, mantendo ou ajustando sua produtividade total. A soja é uma das culturas mais plásticas do agronegócio brasileiro, possuindo habilidade notável de compensar alterações na densidade de plantas através de modificação de seus componentes de rendimento.[2][3][1]

O mecanismo central dessa plasticidade é a regulação da ramificação lateral em resposta à competição intraespecífica por luz, água e nutrientes. Quando uma planta cresce em densidade reduzida, a menor competição por radiação fotossinteticamente ativa (PAR) permite que os brotos axilares situados ao longo do caule principal se desenvolvam em ramos estruturalmente maiores e mais produtivos. Inversamente, em alta densidade, a competição por luz estimula o sombreamento relativo dos ramos internos, reduzindo drasticamente seu número de nós reprodutivos férteis.[4][5][1][2]
A estrutura hierárquica da soja organiza-se em níveis de contribuição produtiva: caule principal (maior potencial, 60-75% das vagens) > ramos de primeira ordem (20-30%) > ramos de segunda ordem (5-10%). Cada nó representa um ponto potencial de inserção de vagens, e a qualidade produtiva de cada nó depende tanto de sua hierarquia quanto das condições ambientais durante o período crítico de floração (R1 a R5).[6][7][8][9]
O número de nós reprodutivos por planta é determinado principalmente durante a fase vegetativa, entre V4 e V6, criando uma “janela crítica” onde decisões de manejo devem ser tomadas. Estudos demonstram que plantas cultivadas em densidade reduzida (200-240 mil plantas/ha) podem emitir 8-10 ramos por planta, enquanto plantas em densidade alta (>350 mil/ha) emitem apenas 2-4 ramos. Contudo—e este é o ponto crucial—essa redução no número de ramos é completamente compensada pelo aumento significativo em vagens por planta no caule principal e ramos remanescentes.[3][10][11][2][6]
Fatores Limitantes à Ramificação Produtiva

Luz e Competição Intra-específica
A competição por luz é o fator ambiental mais determinante para a ramificação lateral em soja. Quando a população ultrapassa 300-320 mil plantas/ha em espaçamento reduzido (<0,45 m entre linhas), ocorre um processo denominado “efeito de sombreamento de ramos internos”.[1][2][4][5]
As folhas posicionadas no topo da planta interceptam preferencialmente a radiação PAR, deixando os ramos em posições mais baixas sob condições de luz vermelha enriquecida (R:FR > 1,0), que sinaliza competição. Este sinal hormonal reduz a síntese de citocininas nos ramos internos, limitando seu crescimento e desenvolvimento de nós reprodutivos. A citocinina é o fitormônio responsável pela divisão celular em meristemas laterais; sua redução por sinal de sombreamento marca o ponto de decisão evolutivo entre investir em estruturas laterais versus crescimento vertical para competir por luz.[5][1]
Os dados de campo demonstram que a cada incremento de 50 mil plantas/ha acima da densidade ótima para determinada cultivar, ocorre redução de 5-8% no número de ramos produtivos. Um estudo comparativo mostrou que em 135 mil plantas/ha, as plantas emitiram ~9 ramos por planta, enquanto em 440 mil plantas/ha, esse número caiu para ~2,5 ramos. Porém—e este é o diferencial da plasticidade—essa redução foi completamente compensada pelo aumento em vagens por planta, mantendo a produtividade de campo relativamente estável.[2][3]
Nutrição Mineral: Nitrogênio, Potássio e Boro
A nutrição mineral exerce controle fino sobre a arquitetura de ramificação. Deficiência de nitrogênio resulta em ramos reduzidos e com menor número de nós reprodutivos, enquanto excesso de nitrogênio promove crescimento vegetativo intenso mas com redução significativa de vagens por nó. A regulação hormonal do nitrogênio ocorre principalmente através da concentração interna de citocininas livres; plantas com N adequado mantêm níveis altos de citocininas, enquanto plantas com déficit de N reduzem a síntese deste hormônio.[12][13]
O potássio é essencial para o transporte eficiente de fotoassimilados dos tecidos-fonte (folhas) para os tecidos-dreno (ramos, vagens). Deficiência de K reduz a alocação de biomassa para ramos laterais, resultando em menor número de vagens mesmo quando o número de ramos é adequado. A absorção máxima de K ocorre entre R2 e R3, exigindo que o elemento esteja disponível em concentrações adequadas no solo (>100 ppm de K trocável) antes desse período.[14][12]
O boro é o micronutriente com maior impacto direto na diferenciação de gemas laterais e frutificação. Sua deficiência durante V4-V6 resulta em morte de gemas laterais e redução drástica de floração nos ramos remanescentes. Estudos com aplicações foliares de boro entre V6 e R1 demonstraram aumento de 17,6% no peso de sementes por planta, principalmente através do aumento em vagens nos ramos laterais. O boro participa da síntese de parede celular e diferenciação meristemática—processos fundamentais na formação de estruturas reprodutivas.[15][16][17][12]
Água: Veranicos Críticos e Estresse Hídrico
A disponibilidade de água entre V6 e V8 é crítica para a sobrevivência de gemas laterais. Um veranico de apenas 7 dias durante V6-V8 causa morte de meristemas axilares e redução de até 8-9% no número de ramos por planta.
Posteriormente, durante R1-R5 (fixação de vagens), períodos de seca induzem aborto de flores principalmente nas posições secundárias dos ramos, onde a competição por carboidratos é maior.[18][19][20][21]
Pesquisas recentes demonstram que seca durante V6 causa redução de 22% na produção de grãos por planta, enquanto seca durante R3 causa redução de 21%—impactos similares nos dois períodos. Contudo, a combinação de seca com deficiência nutricional amplifica drasticamente a morte de gemas e o aborto de flores, criando efeitos sinérgicos negativos.[19][18]
Estratégias de Otimização da Arquitetura
Seleção de Cultivar com Potencial de Engalhamento
A primeira decisão estratégica é a escolha de cultivar compatível com manejo de densidade otimizado. Cultivares modernas indeterminadas diferem geneticamente em seu “potencial de engalhamento”—a capacidade máxima de emitir ramos laterais sob condições não-limitantes. Neste contexto, o Grupo de Maturação Relativa (GMR) funciona como indicador de ciclo, enquanto a arquitetura genética (determinada vs. indeterminada, compacta vs. aberta) indica potencial de ramificação. Pesquisas publicadas na Frontiers in Plant Science confirmam a base genética dessas diferenças arquitetônicas entre cultivares.[4][1]
Aplicação de Bioativos Entre V2 e V5
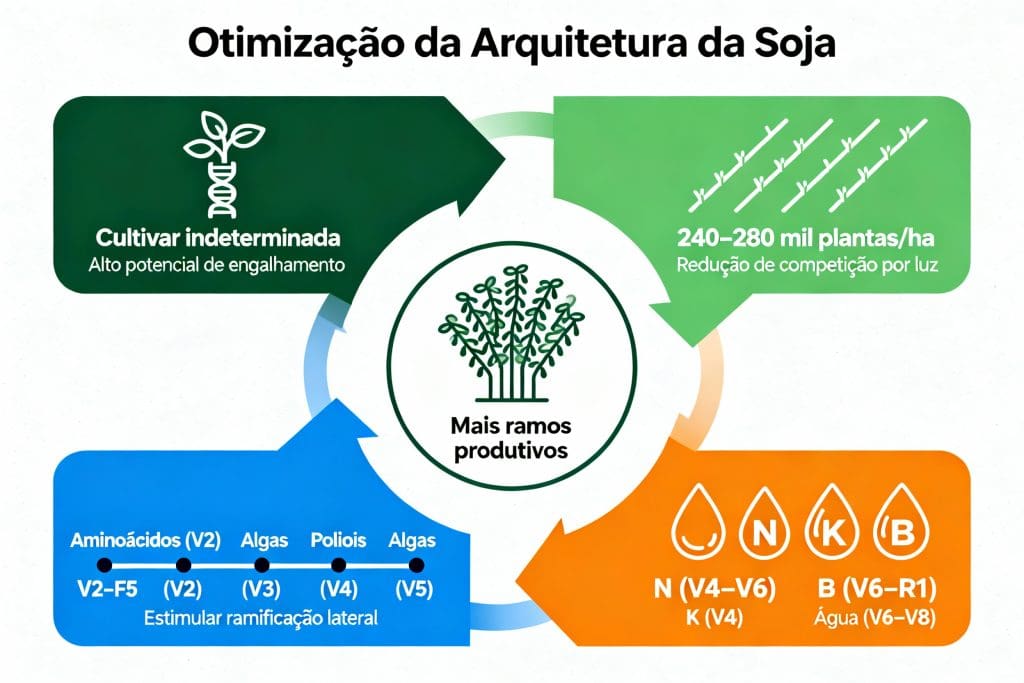
A aplicação estratégica de reguladores vegetais entre V2 e V5 é ferramenta eficaz para potencializar ramificação lateral. Os bioativos dividem-se em compostos orgânicos naturais (derivados de algas, extratos vegetais, aminoácidos) e citocininas sintéticas (benziladenina).
Inoculação com Azospirillum brasilense
Plantas coinoculadas com Bradyrhizobium e Azospirillum apresentam síntese aumentada de fitormônios pelas bactérias, resultando em maior ramificação lateral. A Azospirillum coloniza raízes e aumenta volume explorado do solo, melhorando absorção de água durante V4-V6—período crítico para diferenciação de gemas. Segundo a Embrapa Soja, a coinoculação já é prática recomendada para a cultura e apresenta retorno consistente em ensaios de campo em todo o Brasil.[29]
Manejo de Densidade de Semeadura
O ajuste de densidade é ferramenta com alto retorno potencial quando integrada com bioativos e nutrição. Estudos comparando diferentes densidades demonstram que densidades reduzidas (200-260 mil plantas/ha) resultam em produtividades iguais ou superiores às densidades altas (>300 mil/ha), desde que a cultivar tenha potencial genético de ramificação, condições de água sejam adequadas, nutrição mineral seja balanceada e reguladores de crescimento sejam aplicados estrategicamente. Um estudo publicado em Plant and Soil documenta como a densidade modifica o padrão de desenvolvimento de ramos em diferentes genótipos.[2]
Ferramentas de Diagnóstico
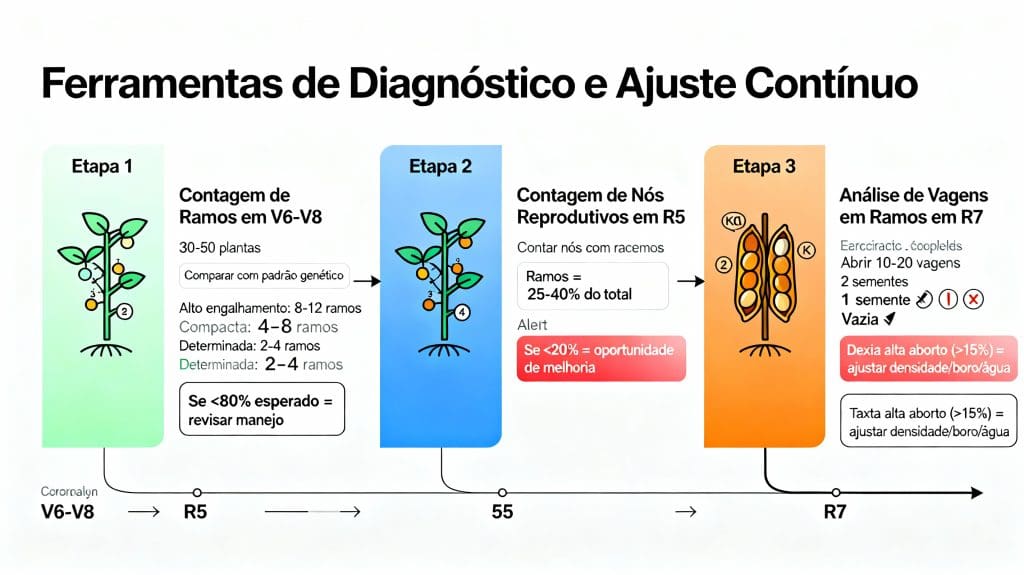
Realize caminhamento no talhão e conte número de ramos por planta em 30-50 plantas durante V6-V8. Compare com expectativa genética: cultivares de alto engalhamento (8-12 ramos), compactas modernas (4-8 ramos), determinadas (2-4 ramos).
Conclusão
A arquitetura de planta é um atributo gerenciável através da combinação de genótipo, densidade de semeadura, biologia do solo, nutrição, bioativos e água. A sequência integrada recomendada: (1) selecione cultivar indeterminada com alto potencial de ramificação; (2) aplique bioativos naturais entre V2 e V5; (3) reduza densidade para 240-260 mil plantas/ha; (4) sincronize cobertura nitrogenada; (5) aplique boro parcelado; (6) garanta água em V6-V8; (7) monitore ramificação em V8.




0 comentários