

O sistema antioxidante enzimático das plantas é o mecanismo que determina quanto tempo o Fotossistema II resiste ao estresse abiótico antes de colapsar. No artigo anterior desta série, vimos que o Fv/Fm cai antes de qualquer sintoma foliar aparecer. A queda reflete a formação acelerada de espécies reativas de oxigênio (EROs) dentro do cloroplasto. O que ainda não discutimos é o que acontece no intervalo entre a formação dessas moléculas e o colapso do Fotossistema II.
Esse intervalo é onde a planta tenta se defender. É também onde a maioria das lavouras perde a batalha produtiva. Noctor e Foyer (1998), em revisão que ainda orienta a área, descreveram a via ascorbato-glutationa como o eixo central da defesa antioxidante em plantas — um sistema enzimático sofisticado, compartimentalizado e dependente de cofatores específicos para funcionar sob pressão ambiental intensa.
O consultor que entende essa fisiologia passa a fazer uma pergunta diferente na lavoura. Não mais "tem sintoma?", mas "esse metabolismo tem o que precisa para aguentar os próximos dez dias de calor?". A resposta depende de três enzimas e de uma cadeia de cofatores que o estresse consome mais rápido do que a planta consegue repor. É o primeiro de uma série de três artigos sobre fisiologia do estresse abiótico publicada no Agrotécnico.
A primeira linha: superóxido dismutase (SOD)
Quando EROs começam a se formar nos tilacoides, a superóxido dismutase (SOD) é a primeira enzima a entrar em ação. Ela catalisa a conversão do ânion superóxido (O₂•⁻) em peróxido de hidrogênio (H₂O₂) e oxigênio molecular. A reação é específica, rápida e localizada nos compartimentos onde o transporte de elétrons acontece: cloroplastos, mitocôndrias e peroxissomos.
A SOD em plantas não é uma enzima única. Barbosa et al. (2014), na Ciência Rural, descreveram três classes de isoenzimas diferenciadas pelo metal cofator: MnSOD nas mitocôndrias, FeSOD nos cloroplastos e Cu/ZnSOD no citosol e nas organelas. Cada isoforma responde a estímulos ligeiramente diferentes. Isso tem uma implicação prática direta: a planta precisa de manganês, ferro, cobre e zinco disponíveis na forma e quantidade corretas para que o sistema funcione com plena capacidade.
Rosa et al. (2020), na Pesquisa Agropecuária Brasileira, demonstraram em soja que linhagens com maior atividade de SOD durante o déficit hídrico no estádio R3 mantiveram produtividade significativamente superior após a reirrigação. A diferença entre as linhagens não estava na intensidade do estresse sofrido. Estava no ritmo com que cada uma conseguia converter o superóxido antes que ele causasse dano oxidativo irreversível.
O comportamento da SOD sob estresse prolongado é previsível. Nos primeiros dias, a expressão dos seus genes se eleva e a atividade enzimática aumenta. Quando o estresse se prolonga por mais de 72 horas em intensidade moderada a severa, a demanda por cofatores metálicos supera a disponibilidade. A atividade começa a cair. A planta continua produzindo superóxido, mas a velocidade de dismutação recua — e o H₂O₂ que deveria ser produto controlado acumula em concentrações problemáticas.
O segundo passo: catalase e ascorbato peroxidase
O peróxido de hidrogênio produzido pela SOD não é inerte. Em concentrações elevadas, o H₂O₂ oxida proteínas, danifica membranas e inativa enzimas do Ciclo de Calvin, incluindo a própria rubisco. O sistema antioxidante tem duas rotas principais para eliminá-lo, cada uma operando em compartimento e condição distintos.
A catalase (CAT) atua principalmente nos peroxissomos. Gill e Tuteja (2010), em revisão na Plant Physiology and Biochemistry, registraram sua taxa de conversão: até seis milhões de moléculas de H₂O₂ decompostas por minuto, gerando água e oxigênio molecular. É uma enzima de alta capacidade, mas baixa afinidade — funciona bem quando a concentração de H₂O₂ é elevada, como nos peroxissomos onde o produto da fotorrespiração se acumula. Nos cloroplastos, a CAT está ausente.
Noctor e Foyer (1998) detalharam o mecanismo que opera nos cloroplastos: a ascorbato peroxidase (APX). Com alta afinidade pelo substrato mesmo em concentrações baixas de H₂O₂, a APX usa o ascorbato como doador de elétrons para reduzir o peróxido em água. Cada molécula de H₂O₂ eliminada consome uma molécula de ascorbato, que passa à forma oxidada (monodehidroascorbato, MDHA). Para que o ciclo continue, o MDHA precisa ser regenerado — processo que consome NADPH e envolve as enzimas dehidroascorbato redutase (DHAR) e glutationa redutase (GR), com glutationa como intermediário.
Esse ciclo, denominado ciclo ascorbato-glutationa, tem uma consequência raramente discutida no campo. Sua capacidade de eliminar H₂O₂ depende da disponibilidade contínua de ascorbato e glutationa nas formas reduzidas. Quando o estresse se prolonga, a taxa de oxidação desses compostos supera a taxa de regeneração. O pool de ascorbato reduzido cai. A APX perde substrato. O H₂O₂ começa a se acumular nos cloroplastos. O dano à proteína D1 do PSII acelera — e o Fv/Fm escorrega para a zona vermelha. Para compreender esse dano ao PSII em detalhe, leia sobre a otimização da fotossíntese e o papel das enzimas antioxidantes no equilíbrio redox celular.
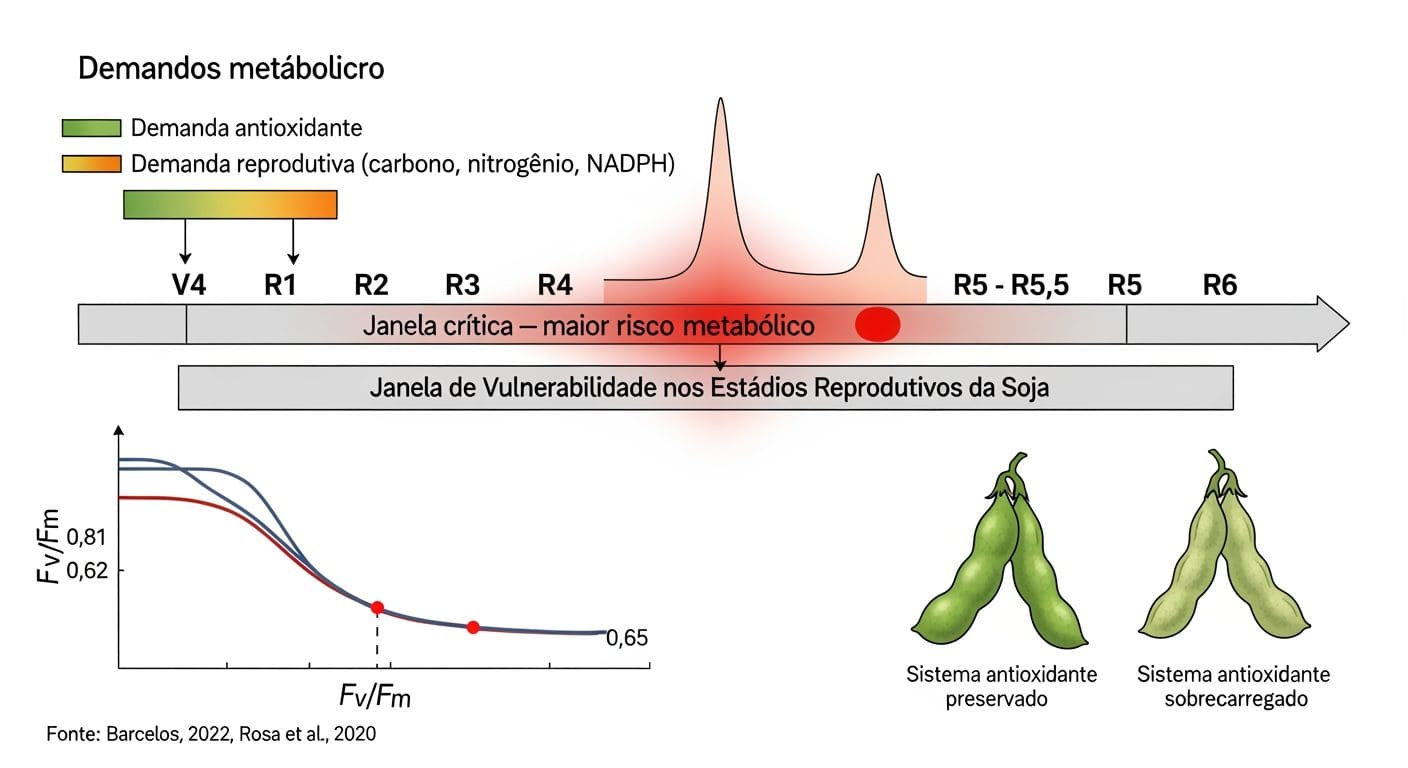
A janela de vulnerabilidade nos estádios reprodutivos
A planta consegue sustentar o sistema antioxidante por períodos curtos de estresse moderado. O colapso começa quando o estresse se prolonga ou coincide com estádios de alta demanda metabólica. Nos estádios reprodutivos, a planta opera com demanda energética máxima — em soja, o florescimento (R1-R2) e o início do enchimento de grãos (R5-R6) são os períodos de maior mobilização de carbono e nitrogênio para as estruturas reprodutivas.
Nessa janela, a disponibilidade de precursores para síntese de enzimas antioxidantes e regeneração dos pools de ascorbato e glutationa é naturalmente menor. Tudo compete pelo mesmo recurso. Barcelos (2022), em dissertação pela USP/ESALQ sobre o efeito da prolina em soja sob déficit hídrico, observou que plantas estressadas no estádio reprodutivo apresentaram queda significativa nos marcadores de atividade antioxidante quando o déficit se prolongou além de cinco dias.
A recuperação após reirrigação foi parcial nos parâmetros enzimáticos. Nos componentes de rendimento, foi incompleta: número de vagens e peso de grãos já comprometidos não se recuperaram com a normalização hídrica. Esse padrão define a janela de vulnerabilidade — o período em que o sistema antioxidante está ativamente consumido, a regeneração dos cofatores compete com a demanda reprodutiva, e qualquer evento de estresse, mesmo moderado, tem potencial de cruzar o limiar que separa a zona amarela da zona vermelha do Fv/Fm.
Sarker e Oba (2018), em estudo publicado na Scientific Reports, confirmaram esse padrão comparando genótipos tolerantes e sensíveis ao déficit hídrico. O genótipo tolerante apresentou incremento de 379% na atividade de APX e 375% na atividade de GR sob estresse severo — contra incrementos muito menores no genótipo sensível. A diferença de produtividade entre os dois foi diretamente proporcional à capacidade de sustentar o ciclo ascorbato-glutationa durante o evento.

A prolina: osmoproteção e antioxidação simultâneas
A prolina ocupa um lugar particular nessa fisiologia porque atua em duas frentes ao mesmo tempo. Como osmoprotetor, ela se acumula no citoplasma e nos cloroplastos sob déficit hídrico, reduzindo o potencial osmótico celular e contribuindo para a manutenção do turgor. Como antioxidante, participa diretamente da eliminação de radicais livres e estabiliza estruturas proteicas e membranares sob pressão oxidativa.
Hare e Cress (1997) descreveram um terceiro papel, menos conhecido: a síntese de prolina consome NADPH, ajudando a manter o Ciclo de Calvin ativo mesmo quando a disponibilidade de CO₂ cai por fechamento estomático. A síntese de prolina funciona como via alternativa de consumo de equivalentes redutores, prevenindo o acúmulo excessivo de elétrons que levaria à maior formação de EROs. O mecanismo é indireto. A consequência para o Fv/Fm é direta.
A síntese de prolina é catalisada pela enzima Δ¹-pirrolina-5-carboxilato sintase (P5CS), cuja expressão depende da disponibilidade de glutamato e de NADPH. Quando o estresse é prolongado, a competição por esses substratos com outras vias metabólicas limita a velocidade de síntese. O sistema protetor é afetado pela mesma pressão que deveria combater (Barcelos, 2022). Para quem trabalha com ajuste osmótico e osmoproteção em plantas, esse mecanismo é o contexto fisiológico que explica quando e por que a prolina exógena tem efeito mensurável.
O que o sistema precisa para não falhar
A interdependência interna do sistema antioxidante é o ponto central desta discussão. Cada componente consome o substrato que outro precisa. A SOD precisa de Mn, Fe, Cu e Zn como cofatores enzimáticos. A CAT depende de ferro heme em seu centro ativo. A APX consome ascorbato continuamente, cuja regeneração demanda NADPH. A glutationa redutase precisa de NADPH e glutationa. A síntese de prolina consome glutamato e NADPH.
O NADPH é produzido pela fase fotossintética que o próprio estresse está comprometendo. Barbosa et al. (2014) sintetizaram esse cenário: quando o estresse quebra o equilíbrio do sistema fotossintético, o cascateamento de deficiências percorre toda a rede antioxidante. Não é uma falha pontual — é um colapso sistêmico que começa nos cofatores e termina no Fv/Fm. Compreender os sinais precoces do estresse oxidativo em campo depende de conhecer essa cadeia.
A planta que chega ao florescimento com alta atividade de SOD, CAT e APX, com pools adequados de ascorbato e glutationa reduzidos e cofatores metálicos disponíveis, tem margem para absorver um evento de calor ou seca moderados sem que o Fv/Fm escorregue para a zona vermelha. A planta que chega ao mesmo estádio com sistema antioxidante já parcialmente consumido por estresses anteriores — mesmo que subclínicos — vai colapsar mais rápido, recuperar mais devagar e produzir menos.
O que isso muda no raciocínio agronômico
O sistema antioxidante enzimático não funciona apenas em situações de emergência. É ativo continuamente, em qualquer nível de pressão ambiental, consumindo cofatores e substratos mesmo sob estresse leve. Intervenções feitas depois que o Fv/Fm já está na zona vermelha atuam sobre um sistema que não tem mais recursos internos para responder.
A lógica preventiva — construir capacidade antioxidante antes das janelas críticas — é a única que pode alterar o resultado produtivo de forma relevante. Isso tem implicação direta sobre quando e como recomendar produtos posicionados para manejo do estresse abiótico. O artigo seguinte desta série discute por que a maioria dessas soluções age na superfície do problema, e o que distingue uma solução de mecanismo de uma solução de alívio imediato. Para quem já acompanhou o debate sobre o aminograma como preditor de eficiência de bioestimulantes, o próximo artigo vai conectar os dois temas.
Gill e Tuteja (2010) sintetizaram essa lógica: a tolerância ao estresse abiótico não depende de uma única enzima ou cofator, mas da capacidade do sistema inteiro de operar de forma coordenada sob pressão contínua. Nenhum componente isolado resolve. O que define o resultado é a condição metabólica da planta no momento em que o estresse chega — e essa condição é construída antes, não durante. Para mais contexto sobre termorregulação e defesa antioxidante em plantas, o Agrotécnico tem material específico.
Conclusão
O sistema antioxidante enzimático — SOD, CAT, APX e o ciclo ascorbato-glutationa — é o mecanismo que determina por quanto tempo o PSII consegue manter o Fv/Fm na zona verde enquanto o estresse abiótico está presente. Não é a ausência de estresse que diferencia as lavouras que produzem das que colapsam. É a capacidade desse sistema de operar de forma coordenada sob pressão contínua.
O colapso começa quando a demanda por cofatores supera a disponibilidade — especialmente nos estádios reprodutivos, quando a competição metabólica entre defesa antioxidante e enchimento de estruturas reprodutivas é máxima. Qualquer evento de estresse moderado nessa janela tem potencial de cruzar o limiar da zona vermelha. O dano ao PSII que se segue não é revertido pela normalização climática. Leia também: fisiologia das plantas e o momento de cada intervenção, bioestimulantes no manejo do déficit hídrico em soja e compostos bioativos e indução de defesa em plantas.
Referências bibliográficas
BARBOSA, M. R. et al. Geração e desintoxicação enzimática de espécies reativas de oxigênio em plantas. Ciência Rural, Santa Maria, v. 44, n. 3, p. 453-460, 2014. Disponível em: https://doi.org/10.1590/S0103-84782014000300011
BARCELOS, G. F. Efeito da utilização de prolina na tolerância ao estresse hídrico em soja. 2022. Dissertação (Mestrado em Fitotecnia) — Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, Piracicaba, 2022.
GILL, S. S.; TUTEJA, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry, v. 48, n. 12, p. 909-930, 2010. Disponível em: https://doi.org/10.1016/j.plaphy.2010.08.016
HARE, P. D.; CRESS, W. A. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regulation, v. 21, n. 2, p. 79-102, 1997. Disponível em: https://doi.org/10.1023/A:1005703923347
NOCTOR, G.; FOYER, C. H. Ascorbate and glutathione: keeping active oxygen under control. Annual Review of Plant Physiology and Plant Molecular Biology, v. 49, p. 249-279, 1998. Disponível em: https://doi.org/10.1146/annurev.arplant.49.1.249
ROSA, V. do R. et al. Estresse hídrico durante a fase reprodutiva de duas linhagens de soja. Pesquisa Agropecuária Brasileira, v. 55, e01736, 2020. Disponível em: https://doi.org/10.1590/S1678-3921.pab2020.v55.01736
SARKER, U.; OBA, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Scientific Reports, v. 8, n. 1, p. 16496, 2018. Disponível em: https://doi.org/10.1038/s41598-018-34944-0
SCIELO BRASIL. Enzymatic antioxidant responses to biostimulants in maize and soybean subjected to drought. Scientia Agricola, 2020. Disponível em: http://www.scielo.br/j/sa/a/W4YB7P3FmCqbp95pJV7Lmrf/
Sobre o autor

Deyvid Rodrigues Bueno
Fundador do Agrotécnico.com.br
Gerente de Produtos · ICL América do Sul · Gestão de Produtos (Business Behavior Institute) · MBA em Gestão, Empreendedorismo e Marketing (PUCRS) · Especialista em Solos e Nutrição de Plantas (FAZU) · Engenheiro Agrônomo (UFMT) · Técnico Agrícola (IFRO)




0 Comentários