

A planta não absorve nutriente porque você colocou adubo no solo. Ela absorve quando consegue. Essa distinção é fundamental para quem quer entender por que a mesma dose de fósforo resulta em respostas tão diferentes entre uma propriedade e outra, ou entre duas épocas de plantio.
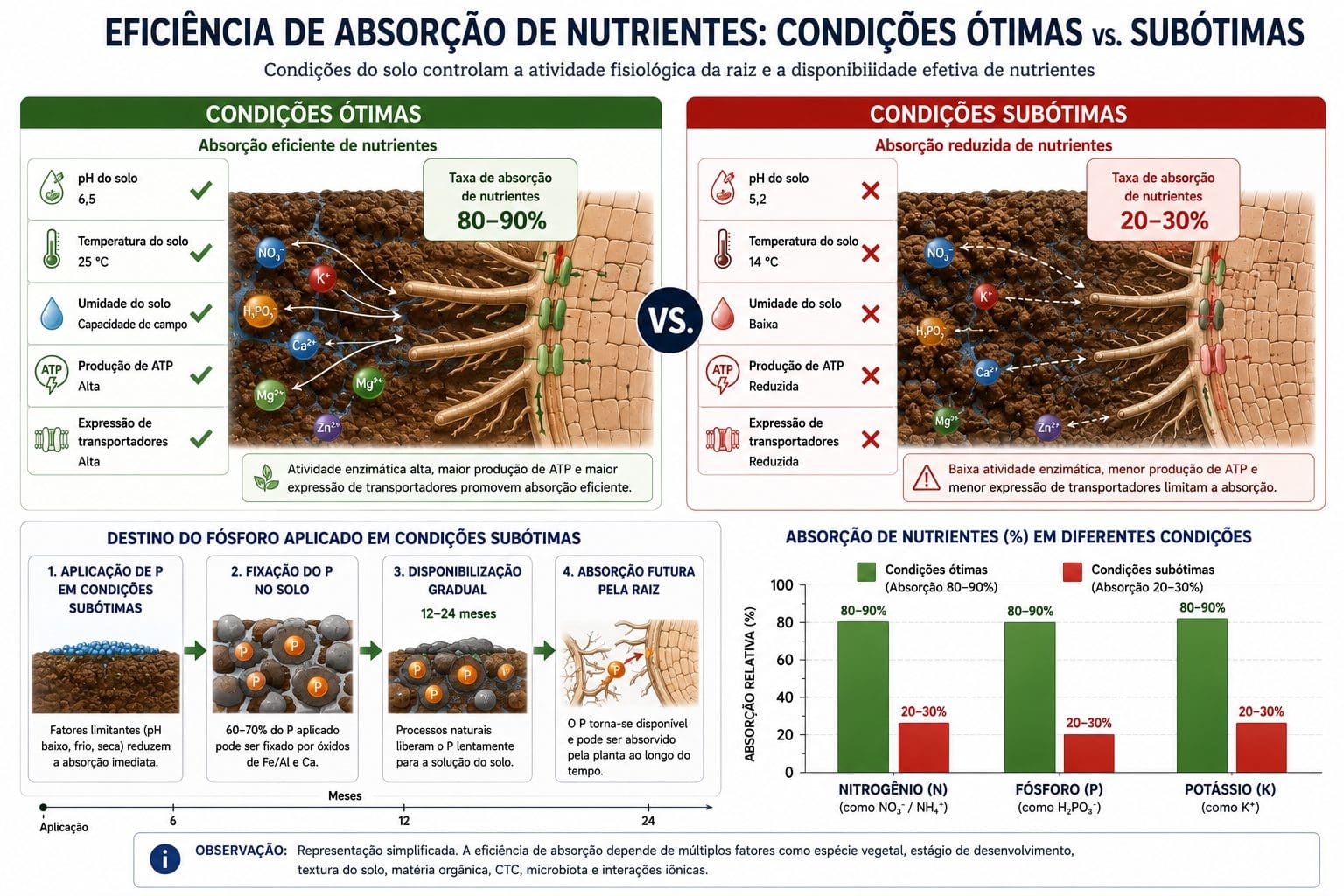
Menos da metade do fósforo solúvel aplicado via adubação é absorvido pela cultura na safra de aplicação em solos tropicais do Cerrado. O fósforo não desaparece. Transforma-se quimicamente (precipita como Al-P ou Fe-P) e permanece no solo como reserva que fica gradualmente disponível nos anos seguintes, conforme a acidez é corrigida. Sousa e Lobato (2004) demonstraram esse desfasamento temporal em Latossolos do Centro-Oeste: a fração fixada de P pode chegar a 60-70% da dose aplicada no primeiro ciclo.
Quando você coloca adubo, coloca uma promessa, não uma garantia. O nutriente precisa estar disponível na solução do solo, na forma iônica correta, em contato com a raiz, atravessando membranas semipermeáveis e, acima de tudo, contar com energia metabólica da própria planta para ser internalizado. Cada um desses filtros é uma barreira que o produtor pode controlar ou deixar negligenciada.
Este artigo percorre os mecanismos de contato íon-raiz, o papel das proteínas transportadoras de membrana, as variáveis que condicionam a absorção e as implicações para o manejo. Você descobrirá por que a absorção de nutriente é um evento fisiológico que depende de física, química e biologia funcionando simultaneamente e por que as condições ótimas raramente ocorrem todas no mesmo dia.
1. Como o nutriente chega à raiz: os três caminhos de transporte no solo
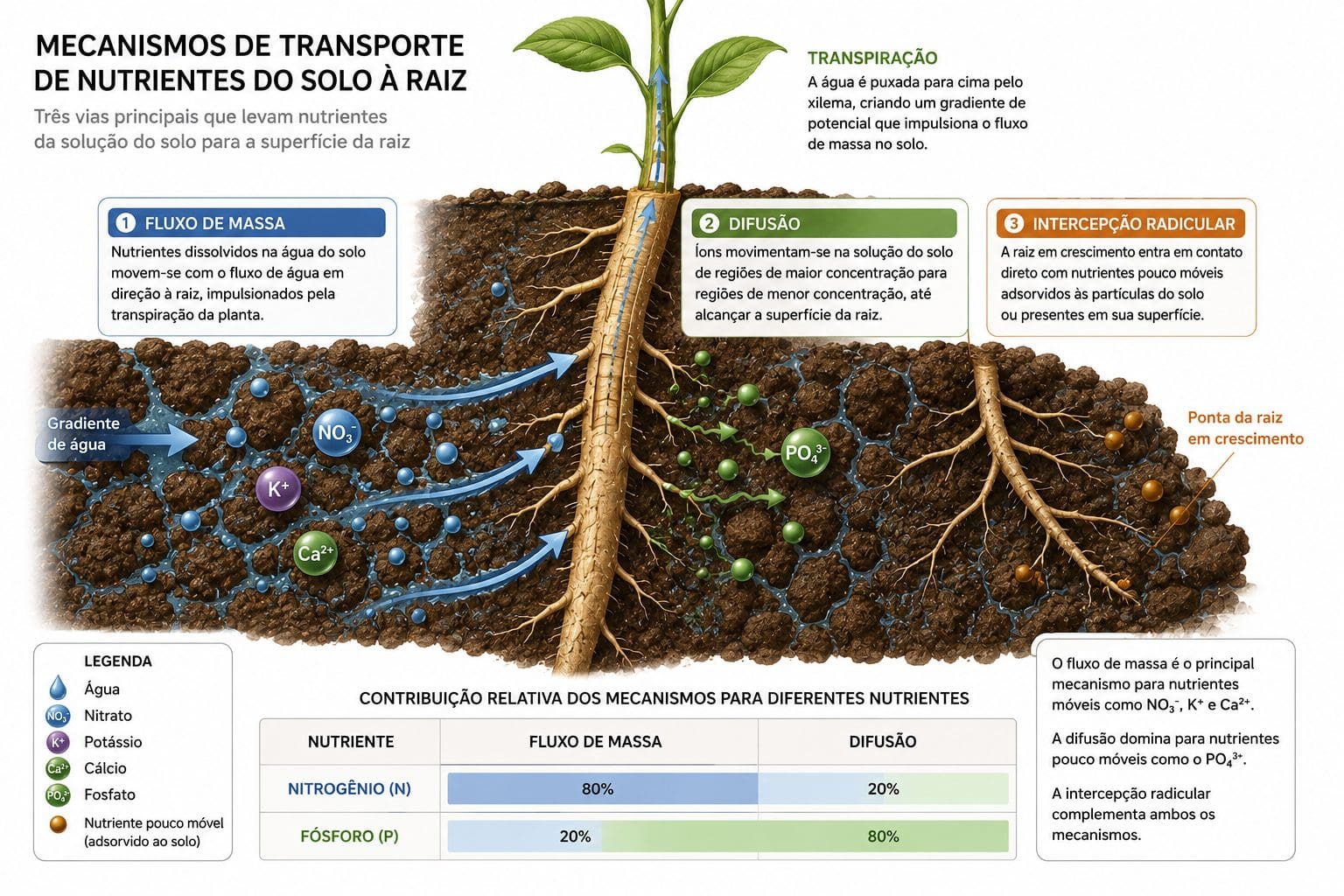
O íon não sai do solo e entra na raiz sozinho. Ele percorre uma distância entre o local onde o fertilizante foi aplicado até a superfície radicular. Barber (1995) descreveu três mecanismos distintos para essa jornada, cada um responsável por uma fração diferente da absorção total conforme o nutriente.
Fluxo de massa é o movimento de íons dissolvidos junto com a água que a planta absorve por transpiração. A raiz puxa água do solo. Dentro dessa água vêm os íons. Nitrato é transportado predominantemente por fluxo de massa. Quando a planta transpira 500 litros de água por metro quadrado em uma safra, ela traz consigo os íons dissolvidos nessa água. Cálcio e magnésio também dependem muito de fluxo de massa, respondendo por 60 a 80% da absorção desses elementos. Potássio é intermediário: 40 a 50% chega por esse caminho.
Difusão é o movimento de íons por gradiente de concentração ao longo de filme de água no solo. Um nutriente em concentração alta próximo à raiz se move para concentração baixa, mais distante. Mas a difusão é lenta. O coeficiente de difusão do fósforo no solo é 10⁻¹² a 10⁻¹⁵ m²/s. Uma molécula de fósforo leva meses para se mover alguns centímetros no solo. Na solução livre, o fósforo se move 10 mil vezes mais rápido. Zinco e cobre têm difusividade ainda menor que fósforo. Esse é o motivo pelo qual o fósforo em um Latossolo se concentra ao redor da raiz muito rapidamente. A demanda ultrapassa a velocidade de reabastecimento. Fósforo é absorvido em 80 a 90% por difusão. Micronutrientes de baixa mobilidade (B, Zn, Cu) também dependem predominantemente de difusão.
Interceptação radicular é o contato direto da raiz com o nutriente durante o crescimento radicular. Conforme a raiz cresce, encontra nutrientes novos. Mas a raiz cresce lentamente. Centímetros por dia. A interceptação contribui com menos de 1% da absorção total para maioria dos nutrientes. É relevante apenas em solos com teores muito altos, ou em casos de raiz pivotante acessando camadas profundas.
A participação de cada mecanismo varia dramaticamente por nutriente. Nitrato chega à raiz em 80 a 85% por fluxo de massa. Fósforo, que é pouco móvel, chega em 10 a 20% por fluxo de massa e 80 a 90% por difusão. Quando o solo está seco, o fluxo de massa diminui drasticamente em dias. O resultado: absorção de nitrato cai 50% em uma semana sem chuva. A absorção de fósforo cai ainda mais porque a difusão também reduz.
A umidade do solo não é um detalhe operacional. É uma pré-condição da absorção.
Para aprofundar em nutrição mineral e fisiologia da absorção, consulte Nutrição Mineral e Fisiologia da Absorção: Fundamentos Científicos para Otimização Nutricional e Arquitetura Radicular e Eficiência na Absorção de Nutrientes.
2. Transportadores de membrana: a porta que precisa estar aberta
O nutriente chegou à superfície da raiz. Agora precisa entrar na célula. Marschner (2012) classifica esse movimento em dois grupos: transporte passivo (raro em macronutrientes) e transporte ativo (dominante).
A membrana plasmática é uma barreira lipídica impermeável a íons. Nenhum íon sai do solo e entra na célula por difusão simples através de lipídios. Cada nutriente precisa de uma proteína transportadora específica. Nitrato entra por proteínas chamadas NRT1 e NRT2. Fósforo entra por proteínas chamadas PHT. Ferro entra por IRT. Potássio entra por KUP ou HAK. Zinco entra por ZIP. A planta não pode absorver mais potássio do que consegue transportar pelas portas disponíveis, por muito que o potássio esteja presente na solução do solo.
A maioria dos nutrientes entra por transporte ativo. A célula gasta ATP para abrir essa porta. A proteína transportadora é acoplada a uma H⁺-ATPase plasmática. Essa enzima bombeia prótons para fora da célula, gerando um gradiente eletroquímico. O nutriente segue esse gradiente e entra na célula contra a sua concentração. Sem ATP, sem transporte. Sem transporte, sem absorção. A H⁺-ATPase é, portanto, a força motriz de toda absorção iônica.
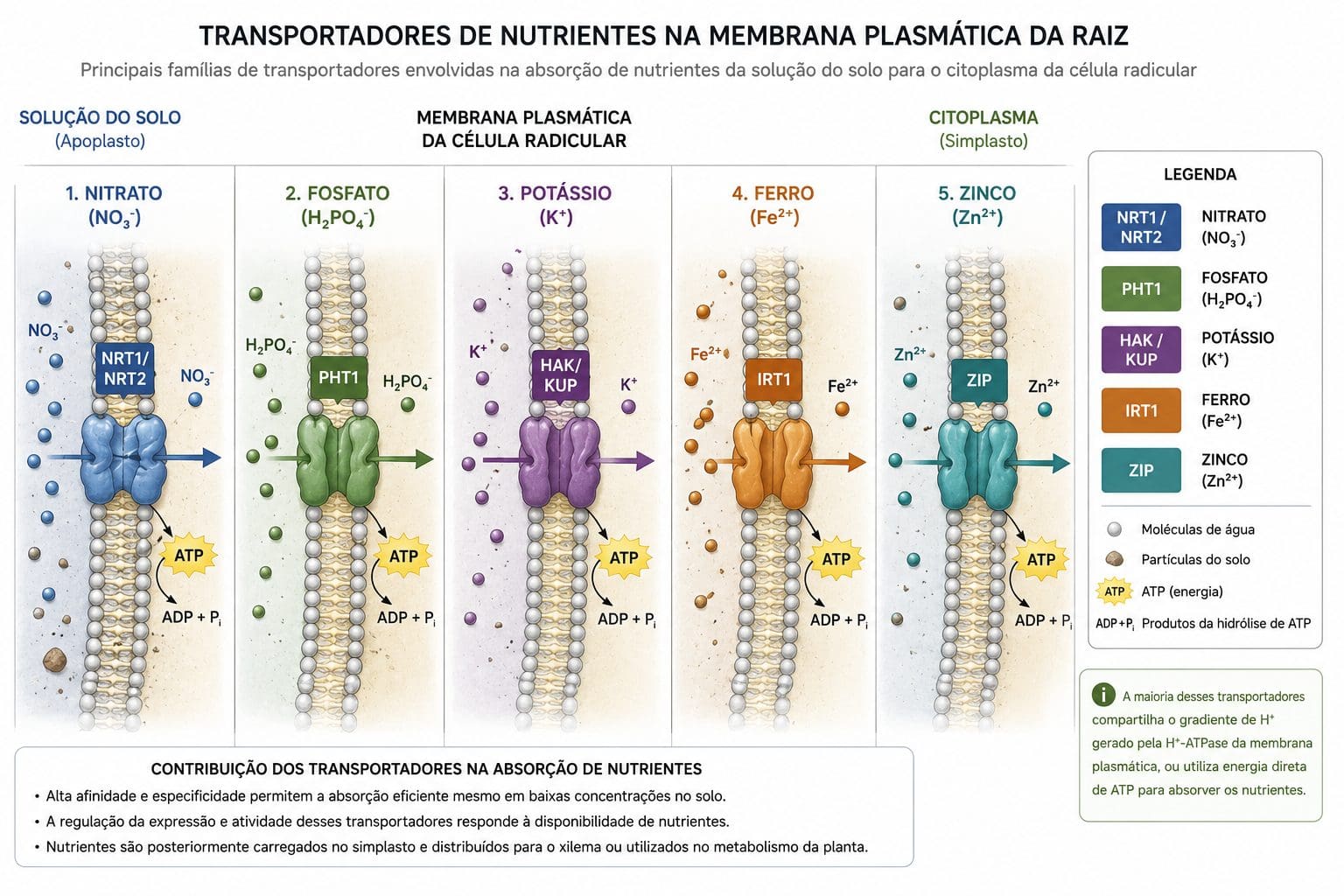
Essas proteínas transportadoras são reguladas de forma sofisticada. Quando o nutriente está em baixa concentração no solo, genes específicos são ativados para aumentar a expressão das proteínas de alta afinidade (HATS — High Affinity Transport System). Essas proteínas conseguem absorver nutrientes em concentrações muito baixas. Quando o nutriente está em excesso, o sistema de alta afinidade é suprimido. A planta economiza energia. Quando a concentração é alta, funciona o LATS (Low Affinity Transport System), menos custoso metabolicamente.
A regulação dessa expressão de transportadores é orquestrada por hormônios vegetais. Sob estresse hídrico, ácido abscísico (ABA) reduz a expressão de transportadores de nutrientes nas raízes para economizar energia. A planta prioriza sobrevivência em vez de absorção. Essa coordenação hormonal é outra camada de controle: mesmo que o nutriente esteja presente, a planta decide se vale a pena investir energia em absorvê-lo naquele momento.
Para entender como os hormônios vegetais coordenam respostas ao estresse, consulte Hormônios Vegetais: Sintético, Precursor e Modulador no Campo e Ácido Salicílico e Suas Aplicações na Fisiologia Vegetal.
3. pH do solo: o regulador central da disponibilidade iônica
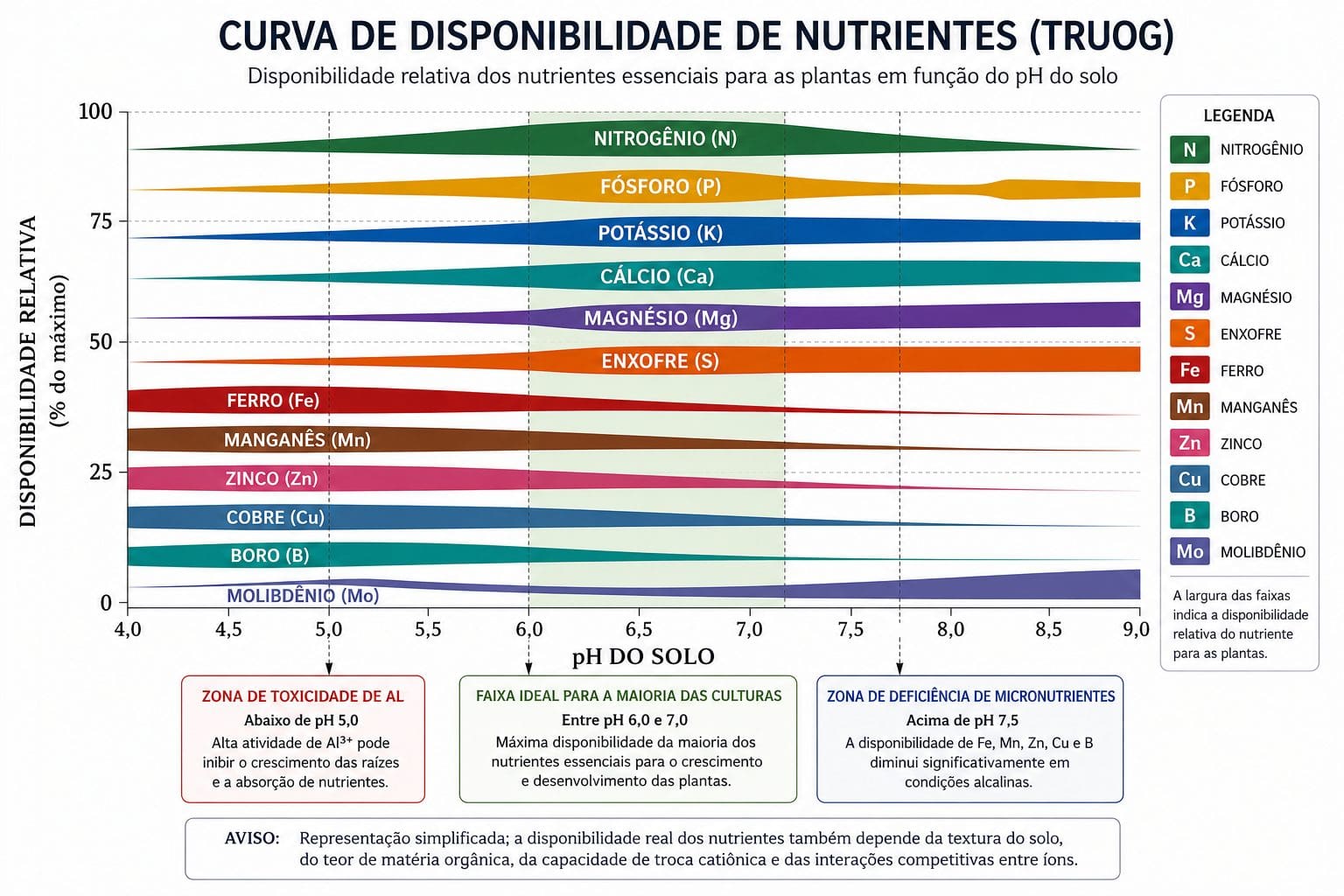
O pH não é apenas um número em um laudo de solo. É o regulador central da especiação química dos íons. O mesmo elemento químico existe em formas iônicas diferentes conforme o pH, cada uma com capacidades radicalmente diferentes de serem absorvidas pelas proteínas transportadoras.
Fósforo é o exemplo mais claro. Entre pH 6,0 e 7,0, o fósforo existe em solução como H₂PO₄⁻. Essa forma é absorvida pela proteína PHT com alta eficiência. Abaixo de pH 5,5, o fósforo se precipita como fosfato de alumínio (Al-P) ou fosfato de ferro (Fe-P). Essas formas não estão em solução. Não estão disponíveis para absorção radicular naquele momento. A faixa ótima de pH para disponibilidade de P é 6,0 a 7,0. Neina (2019) quantificou isso: cada décimo de pH abaixo de 6,0 reduz a disponibilidade de P em média 20% em Latossolos brasileiros.
Micronutrientes como ferro, manganês, zinco e cobre têm disponibilidade máxima em pH baixo (5,0 a 6,5). À medida que o pH aumenta, esses íons precipitam como hidróxidos e carbonatos, tornando-se indisponíveis. Em pH maior que 7,0, a disponibilidade de zinco cai 100 vezes por unidade de pH. Essa é a razão pela qual plantas em solos alcalinos apresentam clorose férrica. O ferro está no solo, mas precipitado como óxido férrico. A raiz não consegue absorver em quantidade ou velocidade suficiente.
Molibdênio tem comportamento inverso. Sua disponibilidade aumenta com o pH. Em solos muito ácidos (pH < 5,0), a deficiência de molibdênio é comum.
Alumínio trivalente é extremamente tóxico em pH abaixo de 5,0. Reduz drasticamente o crescimento radicular e bloqueia diretamente os sítios de absorção de cálcio. A calagem para elevar o pH de 5,0 para 6,0 não é só para adicionar cálcio e magnésio. É principalmente para precipitar o alumínio em óxido (Al(OH)₃) e liberar fósforo fixado em Al-P, tornando-o gradualmente disponível.
Para aprofundar em manejo de rizosfera e correção do solo, leia Rizosfera e Produtividade Agrícola e Como a Relação Ca:Mg Influencia na Compactação dos Solos.
4. Umidade do solo: sem água, o transporte não ocorre
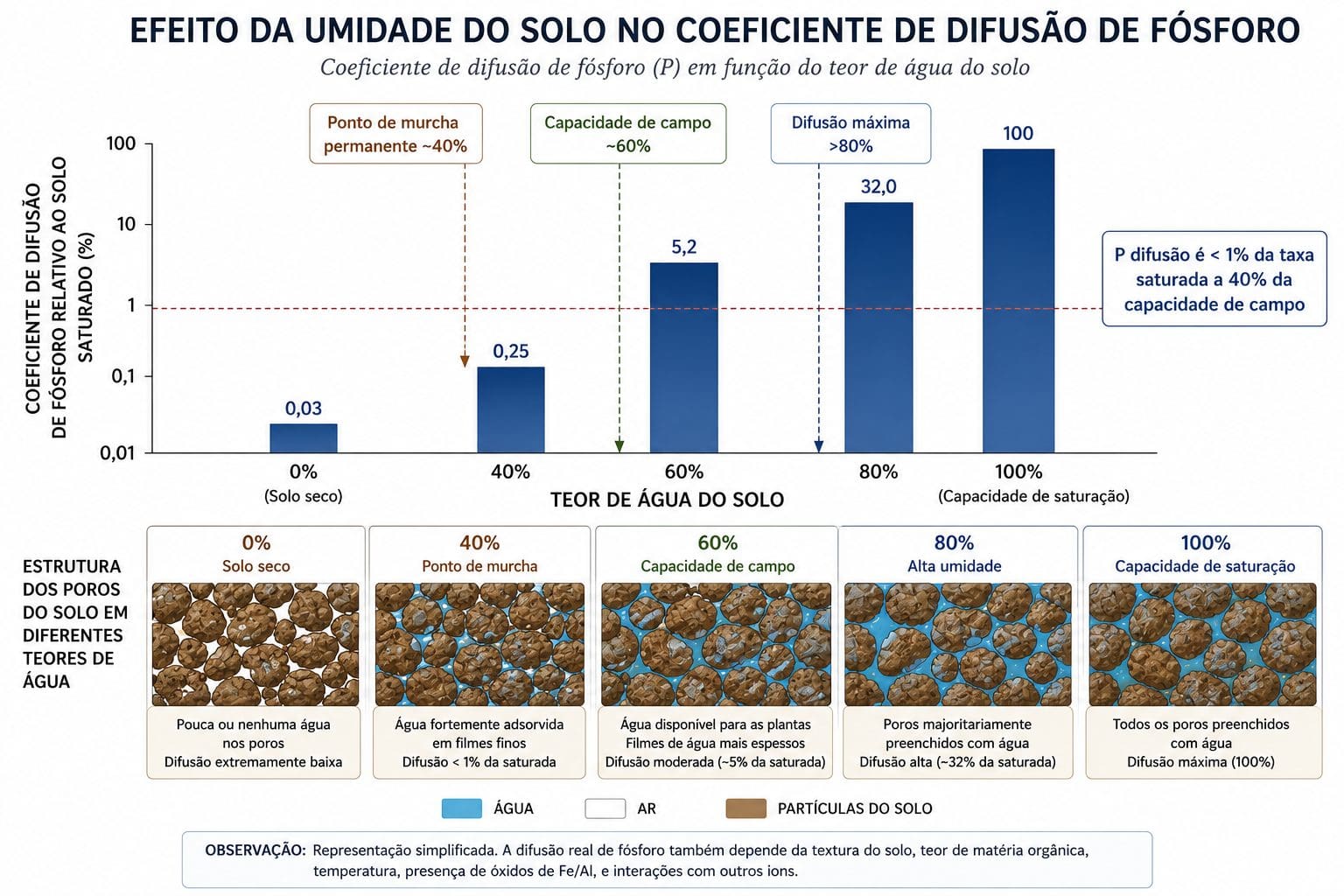
A água no solo não é apenas o recurso da planta. É o veículo de todos os nutrientes. Sem filmes de água contínuos, fluxo de massa para completamente. Difusão desacelera exponencialmente.
Jaleel et al. (2020) mediram o coeficiente de difusão de fósforo sob diferentes umidades. Em solo saturado, o fósforo tem uma velocidade de difusão. Em solo com 60% da capacidade de campo, a difusão se reduz para menos de 1% da difusão em saturação. O caminho do íon através do solo depende de água conectando os poros. Sem água, o caminho desaparece.
A concentração de fósforo na interface raiz-solo depende de quanto fósforo há no solo mais quanto tempo ele teve para se difundir até a raiz. A raiz demanda fósforo continuamente enquanto cresce. Se a difusão não consegue reabastecer a raiz, a concentração local cai para níveis que inibem a absorção. Em solo seco, a raiz fica faminta de fósforo mesmo que o teor total de P no solo seja considerado adequado pelo laudo.
Pesquisas documentam redução de 50 a 60% na absorção de fósforo sob déficit hídrico moderado. Nitratos sofrem redução menor (20 a 30%) porque dependem mais de fluxo de massa, mantido parcialmente mesmo em seca leve. Fósforo sofre redução maior porque depende quase integralmente de difusão, sabotada pela falta de água.
O efeito é pior para nutrientes de baixa mobilidade (P, Zn, Cu) do que para nutrientes de alta mobilidade (N-NO₃⁻).
Para entender como o déficit hídrico impacta a fisiologia das culturas, consulte Bioestimulantes para Déficit Hídrico em Soja e Osmorregulação em Plantas: Ajuste Osmótico para Produtividade.
5. Temperatura: o metabolismo radicular tem janela operacional
A temperatura do solo afeta diretamente três processos: a viscosidade da água (e portanto a velocidade de difusão de íons), a fluidez das membranas lipídicas das raízes e a atividade catalítica das proteínas transportadoras.
Abaixo de 12°C, a inibição de reações bioquímicas nas raízes reduz drasticamente a absorção de água e nutrientes. Khan et al. (2021) estudaram trigo sob frio: a H⁺-ATPase plasmática trabalha com atividade reduzida em temperaturas baixas. Sem bombeamento de prótons, sem transporte ativo. Micronutrientes como zinco e ferro dependem quase integralmente de transporte ativo. Em solo frio, a absorção de Zn pode cair 80 a 90%.
- Soja: 22 a 28°C de temperatura de solo
- Milho: 25 a 30°C de temperatura de solo
- Café: 20 a 25°C de temperatura de solo
O plantio antecipado de soja no Cerrado (agosto, setembro) enfrenta essa barreira fisiológica. O solo ainda não aqueceu o suficiente. A absorção de micronutrientes é reduzida. Folhas mais velhas ficam amareladas. O produtor pensa em deficiência de fertilizante. Na verdade, é falta de energia nas raízes.
Acima de 35 a 40°C no solo, ocorre o oposto. A concentração de proteínas de absorção de nutrientes nas raízes diminui. Estudo com tomateiro submetido a temperatura de raiz de 39°C mostrou queda de 50% nas proteínas que absorvem nitrogênio e fósforo. O efeito negativo foi maior nas raízes comparado à parte aérea porque o metabolismo radicular é mais sensível.
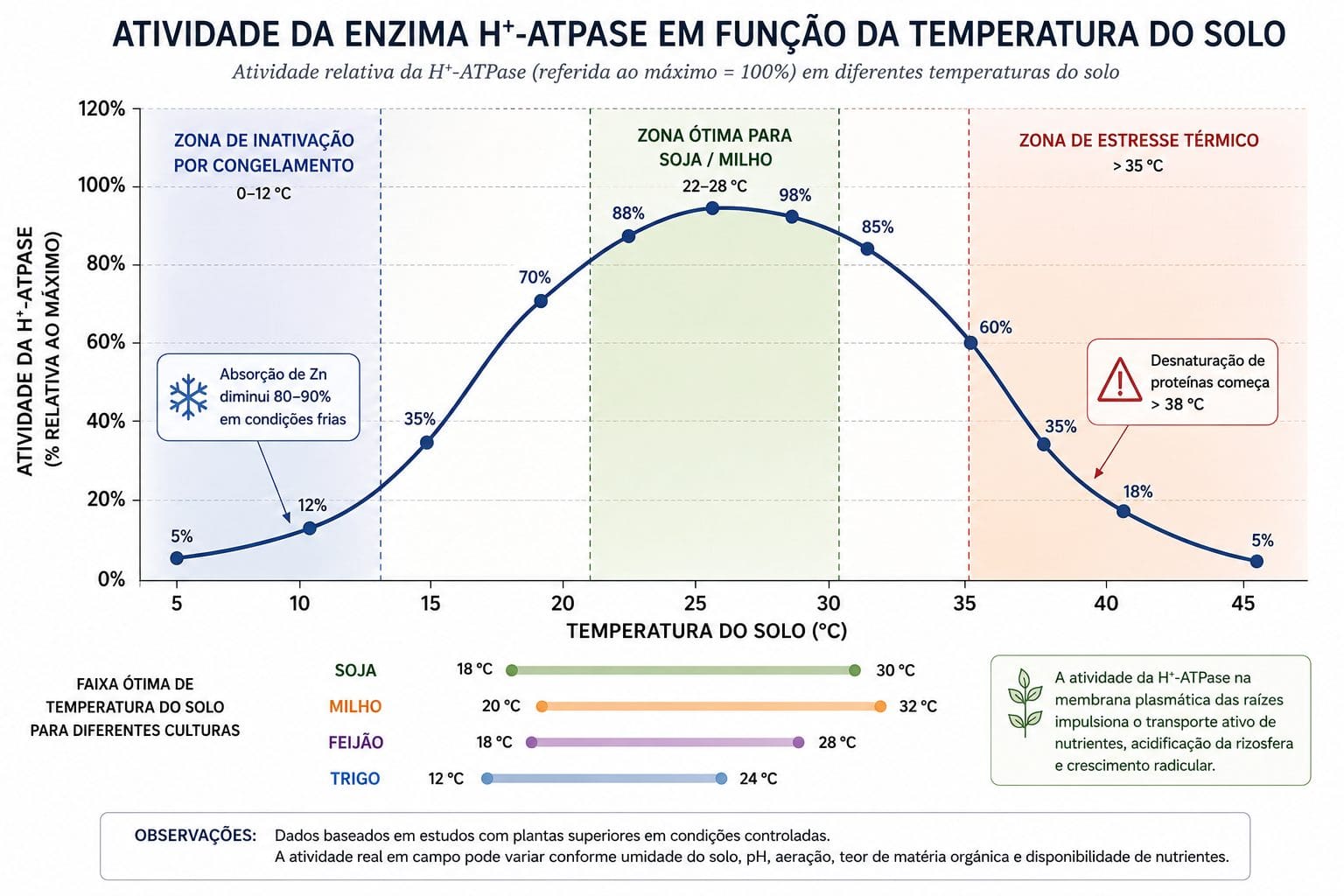
Para entender como a temperatura afeta a fisiologia vegetal, veja Termorregulação em Plantas: Adaptação ao Estresse Térmico e Crioproteção em Plantas: Mecanismos para Baixas Temperaturas.
6. Antagonismos iônicos: quando um nutriente fecha a porta para o outro
Nutrientes competem pelos mesmos sítios de transporte quando têm raio iônico e carga similares. Isso não é defeito do sistema radicular. É a realidade inescapável da bioquímica de membrana. Quando a concentração de um íon no solo está muito alta, ele ocupa mais transportadores. Outros íons similares ficam sem porta de entrada.
O tripé clássico é potássio, cálcio e magnésio. Quando o teor de potássio na solução do solo está elevado, ele compete pelos transportadores com cálcio e magnésio. Adubação excessiva com cloreto de potássio induz deficiência de magnésio visível nas folhas. As folhas mais velhas ficam com verde descolorido e margens castanhas. O magnésio está no solo. Mas o potássio ocupou as portas de entrada.
A relação entre esses três nutrientes no solo importa mais do que o teor absoluto de cada um. A relação Ca:Mg no solo não deve ultrapassar 5:1 para que a absorção de Mg não seja comprometida. Para manter equilíbrio catiônico adequado, recomenda-se que K responda por 2 a 3% da capacidade de troca catiônica (CTC), enquanto Ca + Mg respondam por 70 a 85%.
Fósforo em concentração muito alta no solo pode induzir deficiência de zinco. Ferro em concentração alta reduz a absorção de manganês. Amônia (NH₄⁺) compete vigorosamente com potássio, cálcio e magnésio.
Calagem excessiva em soja no Cerrado elevando a relação Ca:Mg acima de 6:1 resulta em deficiência de magnésio mesmo com teores absolutos de Mg adequados.
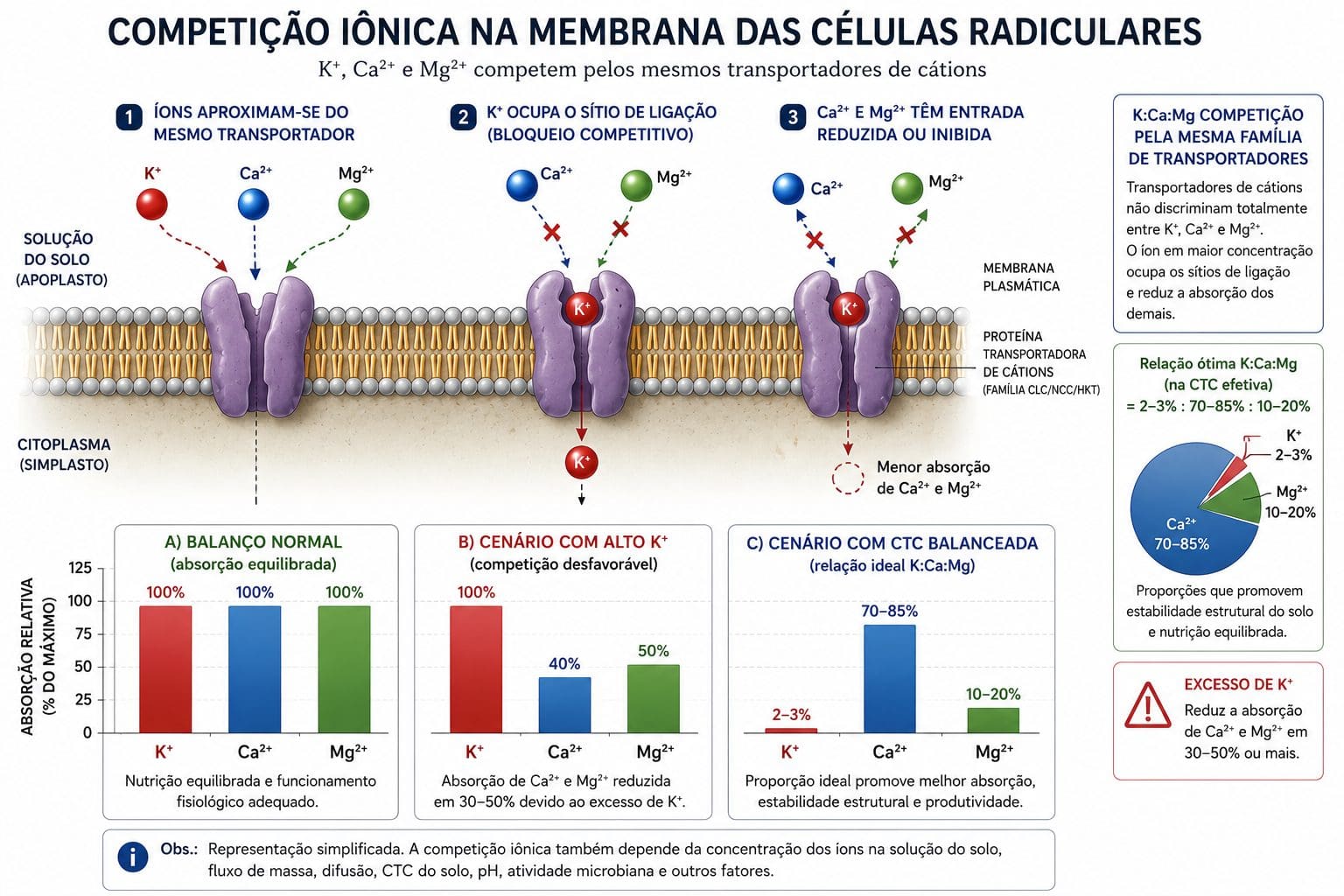
Para aprofundar em equilíbrio catiônico, consulte Como a Relação Ca:Mg Influencia na Compactação dos Solos e Fatores que Interferem na Absorção de Nutrientes pelas Plantas.
7. Fotossíntese e energia: a absorção ativa depende do que acontece na folha
O transporte ativo de nutrientes consome ATP. A maioria do ATP das raízes vem diretamente da fotossíntese foliar ou indiretamente via importação de sacarose fotossintetizada na folha. Hussain et al. (2024) documentaram isso: qualquer condição que reduza a taxa fotossintética afeta diretamente a capacidade da raiz de absorver nutrientes.
Existe um loop de retroalimentação negativo. Déficit hídrico reduz a abertura estomática. Menos CO₂ entra. A fixação fotossintética cai. Menos fotoassimilados são produzidos. Menos glicose chega às raízes via floema. Menos ATP é gerado nas raízes. A H⁺-ATPase trabalha com menos combustível. O gradiente eletroquímico cai. O transporte ativo de nutrientes desacelera. Menos nutrientes são absorvidos.
Doenças foliares como ferrugem asiática reduzem a área foliar ativa e saudável. Menos luz é capturada. Menos ATP é gerado. As raízes sofrem. Excesso de sombreamento reduz a taxa fotossintética. As raízes ficam desabastecidas de energia. Fitotoxicidade por herbicida inibidor de fotossíntese bloqueia a geração de ATP nos cloroplastos. As raízes param de absorver.
O produtor que aplica adubação de cobertura nitrogenada em milho com folhas amareladas por déficit hídrico não resolve a deficiência de N aumentando a dose. A porta de entrada radicular está fechada por falta de energia. O nitrato fica no solo. Aumentar a dose é inútil naquele momento.
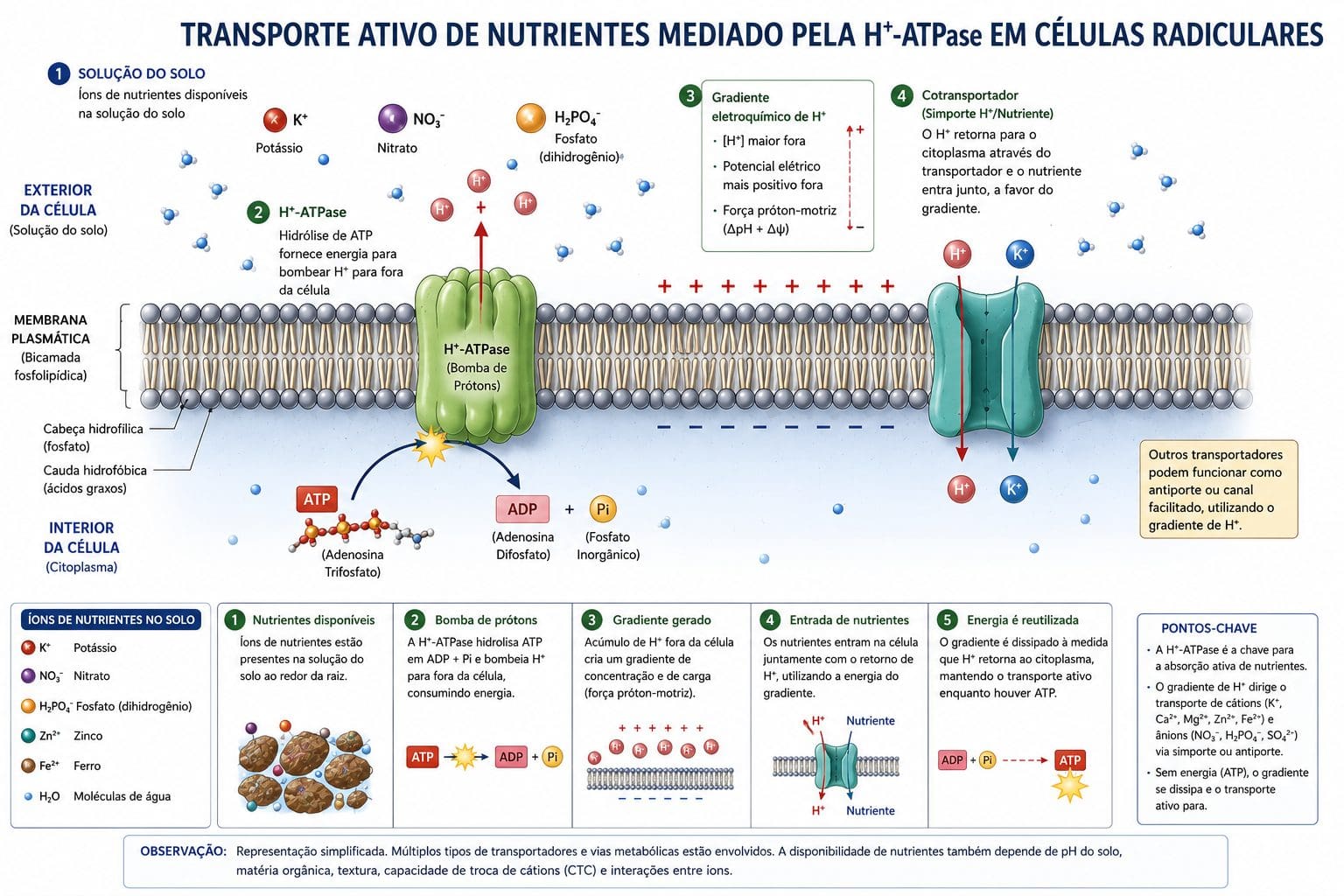
Para aprofundar em fotossíntese e eficiência energética, consulte A Otimização da Fotossíntese e Eficiência Produtiva, Aumentando a Eficiência da Fotossíntese nas Culturas e Fv/Fm: Fluorescência da Clorofila como Indicador de Estresse Abiótico.
8. Microbioma do solo: como os microrganismos estendem o alcance da raiz
A superfície absortiva de uma raiz de soja não-micorrizada é limitada ao comprimento dos pelos radiculares. Fungos micorrízicos arbusculares (FMAs) produzem hifas que se estendem muito além da zona de depleção de nutriente ao redor da raiz. Santos et al. (2012) quantificaram: essas hifas multiplicam a área de solo que a planta consegue explorar em 10 a 100 vezes.
Nutrientes com baixa mobilidade (P, Zn, Cu) são os mais beneficiados pela simbiose. São exatamente os nutrientes que mais dependem de difusão lenta. A hifa fúngica alcança poros do solo onde a raiz não chega. O fungo absorve o nutriente ali e o transporta até o córtex radicular da planta. Em troca, a planta fornece fotoassimilados (açúcares) ao fungo. É uma verdadeira simbiose mutualista.
A associação micorrízica ocorre em 80 a 90% das plantas terrestres. Em soja, a colonização por FMAs é comum em solos não-fumigados. O aumento na absorção de P em plantas bem colonizadas pode atingir 50% em solos com P adequado e até 250% em solos deficientes de P.
Bactérias rizosféricas solubilizadoras de fosfato convertem fósforo inorgânico insolúvel (Al-P, Fe-P, Ca-P) em formas solúveis e disponíveis. Fazem isso via ácidos orgânicos que baixam o pH local e dissolvem as precipitações de P. Espécies como Bacillus e Pseudomonas são comuns em Latossolos brasileiros.
As práticas de manejo agrícola afetam drasticamente a funcionalidade do microbioma. Fungicidas sistêmicos aplicados no sulco de semeadura reduzem a colonização por FMAs nos primeiros 30 a 45 dias após emergência. Nessa fase crítica, a planta tem maior demanda por P para crescimento inicial. Sem FMAs colonizando, a absorção de P cai. Rotação de culturas mantém diversidade funcional. Monocultura reduz essa diversidade.
O manejo do microbioma é silencioso.
Mas é absolutamente crítico para aproveitar o potencial produtivo.
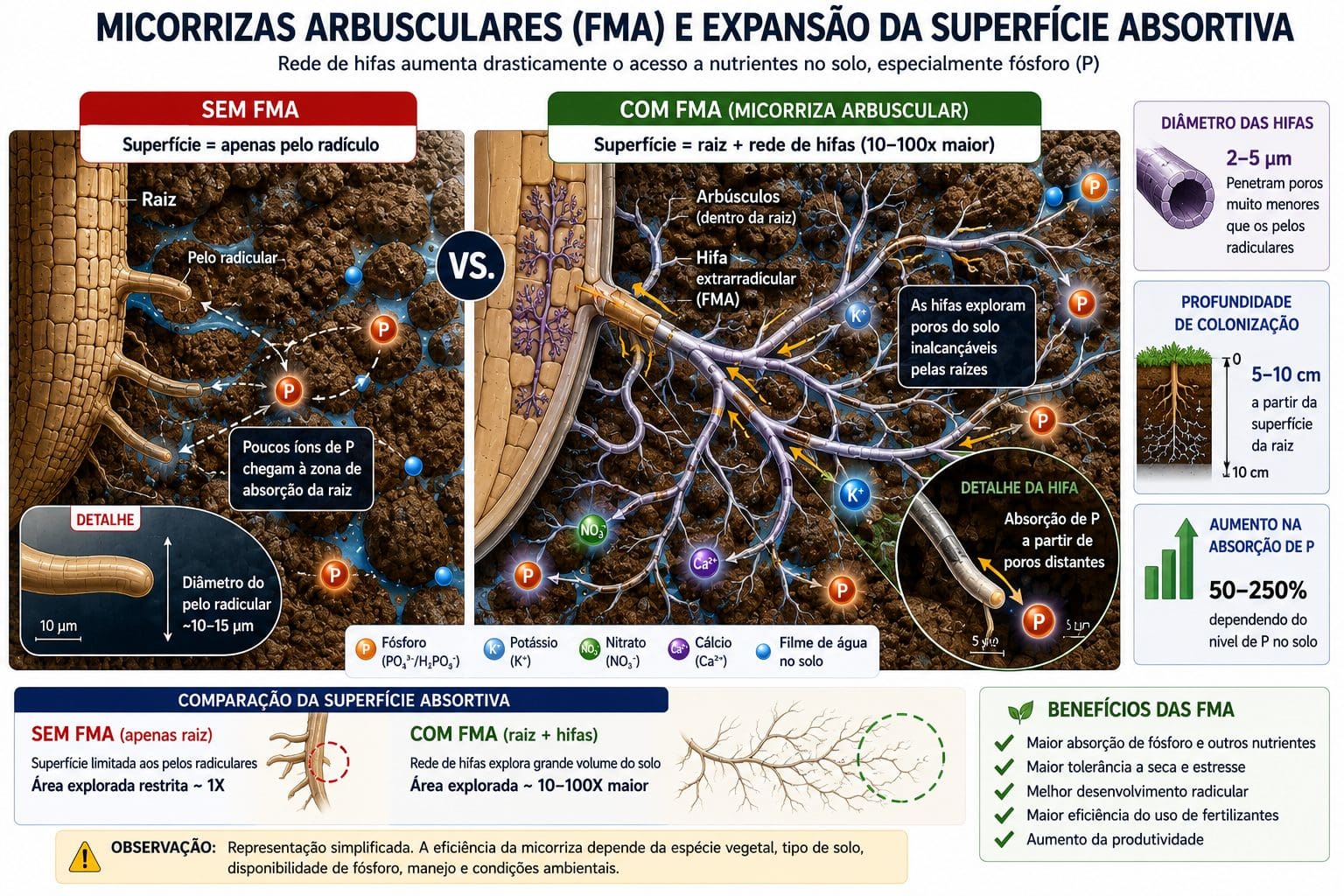
Para entender a dinâmica do microbioma do solo, leia Microbioma do Solo: Sequenciamento Genético e Manejo Racional, Microrganismos e Eficiência no Aproveitamento de Nutrientes e Estresse Abiótico e a Liberação de Exsudatos Radiculares.
9. Implicações práticas: o que muda no manejo quando se entende absorção
A decisão de momento e forma de aplicação precisa considerar se as condições de pH, umidade e temperatura permitem absorção eficiente. Uma adubação fosfatada em solo seco ou muito ácido é dose que aguarda a condição correta para funcionar.
Muitos produtores aplicam fósforo na semeadura em Latossolo com pH 5,2. Pensam que está feito. O fósforo se precipita como Al-P e Fe-P nos dias seguintes. A planta não o absorve naquela safra. Apenas na safra seguinte, após calagem corrigir o pH para 6,0 a 6,5, o P fixado começa a ficar gradualmente disponível. O produtor que entende esse mecanismo faz calagem 18 a 24 meses antes de adubação pesada de P, não simultaneamente.
A adubação foliar não substitui a absorção radicular de macronutrientes em quantidade. Mas contorna limitações edáficas temporárias para micronutrientes de baixa mobilidade (B, Zn, Mn). Aplicação foliar de Zn em soja plantada cedo no Cerrado com solo ainda frio fornece Zn disponível à folha enquanto a raiz não consegue absorver. Depois, quando o solo aquece, a raiz absorve Zn do solo melhor. A deficiência visual desaparece.
A calagem para corrigir pH não é apenas química para adicionar Ca e Mg. É fisiologia: elevar o pH de 5,2 para 6,0 antes de aumentar a dose de P é pré-requisito da absorção. Sem essa correção, o P solúvel aplicado apenas aumenta a fração fixada em Al-P.
Manejo do microbioma pode ser tão impactante quanto adubação. Preservar cobertura de solo, evitar fungicidas de sulco desnecessários e praticar rotação de culturas são práticas que mantêm a atividade de FMAs.
O técnico que analisa apenas o laudo de solo sem considerar o pH atual, a temperatura de solo na data de plantio e a umidade no período de maior demanda perde metade da equação.
Para aprofundar em resiliência metabólica e manejo integrado do estresse, consulte Resiliência Metabólica em Plantas: Estresse Abiótico e Produção e Sistema Antioxidante SOD, CAT e APX no Estresse Abiótico em Plantas.
Conclusão
A absorção de nutrientes é a resultante de uma cadeia integrada de condições: físicas (temperatura, umidade), químicas (pH, disponibilidade iônica, antagonismos) e biológicas (microbioma, fisiologia radicular). Nenhuma condição isolada é suficiente. Todas precisam estar alinhadas dentro de janelas operacionais específicas.
O fertilizante é o insumo. A absorção é o evento fisiológico. Entre os dois existe um conjunto de filtros bioquímicos que a planta não controla, mas o produtor e o técnico podem manejar com precisão. O nutriente no solo que não é absorvido hoje fica como reserva para amanhã. Isso não é desperdício. É fisiologia operando conforme a realidade de campo, onde condições nunca são simultaneamente ideais.
Compreender esse desfasamento temporal entre aplicação física de fertilizante e absorção fisiológica é crítico para decisões de manejo. A maior parte da safra no Cerrado e em regiões tropicais opera em condições subótimas de absorção. Raramente todas as variáveis estão no ótimo no mesmo dia. Esse é um dos motivos pelos quais a produtividade fica abaixo do potencial genético: não é falha na adubação, é fisiologia da absorção limitando a resposta.
A regulação molecular dos transportadores de membrana abre perspectiva futura para desenvolvimento de genótipos com maior eficiência de uso de nutrientes (NUE). Pesquisas em edição gênica via CRISPR estão identificando alelos que conferem maior expressão de transportadores. Cultivares de soja e milho com maior NUE estão em desenvolvimento em universidades brasileiras. Mas enquanto isso não está disponível em larga escala, a fisiologia que você acabou de ler é seu instrumento para maximizar a absorção com o germoplasma que tem agora.
Referências
BARBER, S. A. Soil Nutrient Bioavailability: A Mechanistic Approach. 2. ed. New York: John Wiley & Sons, 1995.
CHE, J.; RICACHENEVSKY, F. K.; DENG, F. Micronutrients movement from soil to the grains: role of plant membrane transporters. Frontiers in Plant Science, v. 13, p. 1179674, 2023. DOI: 10.3389/fpls.2023.1179674
FAQUIN, V. Nutrição Mineral de Plantas. Lavras: UFLA/FAEPE, 2005.
HARTEMINK, A. E.; BARROW, N. J. Soil pH and nutrient availability: an update on the classical approach. Geoderma, v. 425, p. 116071, 2023. DOI: 10.1016/j.geoderma.2023.116071
JALEEL, C. A. et al. Genetic regulation of water and nutrient transport in water stress tolerance in roots. Journal of Experimental Botany, v. 71, n. 1, p. 10–22, 2020.
KHAN, F. et al. Cold stress in wheat: plant acclimation responses and management strategies. Frontiers in Plant Science, v. 12, p. 676884, 2021. DOI: 10.3389/fpls.2021.676884
MARSCHNER, P. (Ed.). Marschner's Mineral Nutrition of Higher Plants. 3. ed. Academic Press, 2012.
NEINA, D. The role of soil pH in plant nutrition and soil remediation. Applied and Environmental Soil Science, v. 2019, p. 5794869, 2019. DOI: 10.1155/2019/5794869
SANTOS, P. M. et al. Micorriza arbuscular e a tolerância das plantas ao estresse. Revista Brasileira de Ciência do Solo, v. 36, n. 6, p. 1743–1783, 2012. DOI: 10.1590/S0100-06832012000600001
SOUSA, D. M. G.; LOBATO, E. (Eds.). Cerrado: Correção do Solo e Adubação. 2. ed. Brasília: Embrapa Informação Tecnológica, 2004.
HUSSAIN, H. A. et al. Impacts of drought on photosynthesis in major food crops. Plants, v. 13, n. 13, p. 1808, 2024. DOI: 10.3390/plants13131808




0 Comentários