

Hormônios vegetais na agricultura vão além da molécula aplicada. Entenda como a capacidade de resposta da planta, precursores e moduladores determinam se ela responde.
Você já viu uma lavoura de soja com boa nutrição, solo manejado, sem estresse visual aparente perder pegamento de vagens sem explicação clara? Ou aplicou um produto posicionado como hormonal no estádio correto e o resultado ficou abaixo do esperado? Essas situações têm algo em comum que raramente aparece na conversa técnica: a variável ignorada não é o produto. É a condição da planta naquele momento.
Durante décadas, a fisiologia vegetal aplicada operou sob uma premissa direta. Mais hormônio equivale a mais resposta. Mais auxina gera mais raiz. Mais giberelina gera mais crescimento. Essa lógica fundamentou os primeiros reguladores vegetais sintéticos e ainda orienta muitas recomendações de campo. O problema é que ela descreve apenas metade da equação.
Trewavas (1982) demonstrou que a resposta da planta depende da sua capacidade de perceber o sinal hormonal, não apenas da concentração do composto externo.
Por volta de 12 anos depois Bradford e Trewavas (1994) formalizaram esse conceito ao mostrar que as curvas de resposta à dose variam com o estado fisiológico, a temperatura e a idade do tecido e que os limiares de ativação mudam conforme a condição da planta naquele momento.
Uma molécula pode estar presente, ser aplicada na dose correta e ainda assim não gerar a resposta esperada se a planta não estiver em condição de responder naquele momento. Esse princípio muda a pergunta técnica central: não é "tem hormônio no produto?", mas "a planta está em condição de responder?".
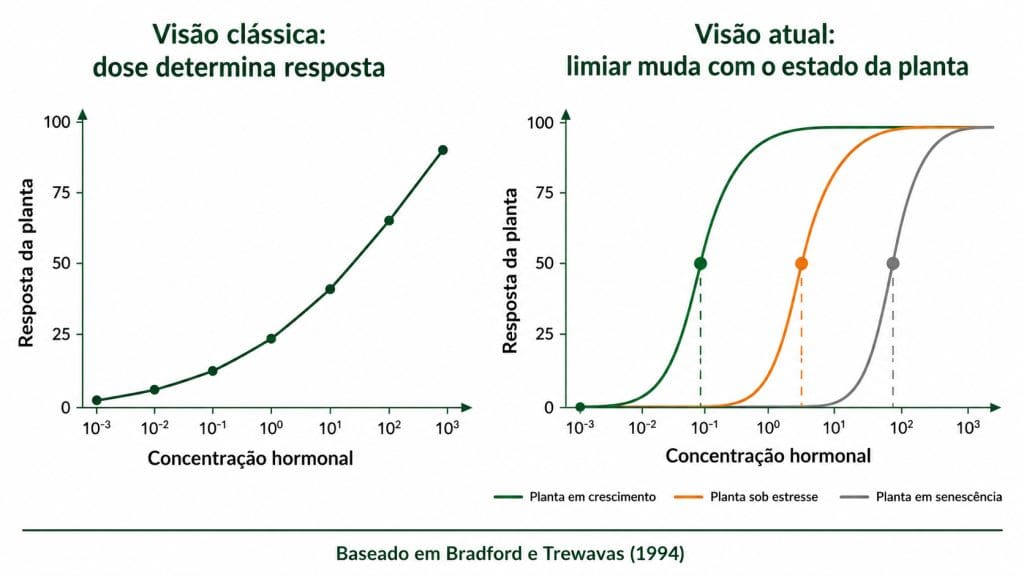
Gráfico 1. Em plântulas de milho com deficiência de zinco, o triptofano se acumula nos tecidos sem ser convertido em IAA ativo. Takaki e Kushizaki (1970) mostraram que a enzima responsável pela conversão requer zinco como cofator estrutural. O precursor estava presente. A rota estava bloqueada.
A planta regula a própria resposta. O agrônomo que entende esse mecanismo decide antes de aplicar, não depois de colher. Neste artigo, percorremos os três tipos de intervenção que o mercado trata como equivalentes, os mecanismos que diferenciam cada um e as perguntas que tornam qualquer recomendação mais precisa. As culturas de referência são soja, milho, café e cana-de-açúcar.
Três coisas que o mercado chama de ação hormonal
Quando um produto é posicionado como hormonal, três mecanismos completamente distintos podem estar sendo descritos. Tratá-los como equivalentes é onde a recomendação começa a falhar.
O hormônio sintético é uma molécula desenvolvida para interferir diretamente em rotas de crescimento vegetal. Trinexapac-etil, cloreto de mepiquate e etefom são exemplos com mecanismo de ação bem documentado, eficácia comprovada em condições específicas e janela de aplicação estreita. São ferramentas de precisão. Funcionam dentro da especificação de estádio, dose e condição da planta. Fora dela, o efeito pode ser diferente do esperado.
O precursor hormonal é um aminoácido ou metabólito que a planta pode usar para sintetizar hormônio endógeno. O triptofano, precursor da auxina ativa pela via do ácido indol-3-pirúvico (IPyA), é o exemplo mais citado no mercado de bioestimulantes. A palavra central é "pode". O precursor oferece um caminho metabólico possível, não uma resposta garantida. A conversão depende de enzimas ativas, cofatores disponíveis e de a planta ter demanda de crescimento naquele momento.
O modulador fisiológico não é hormônio nem precursor direto. Ele favorece rotas endógenas da planta, melhora o equilíbrio oxidativo, reduz dano por estresse e influencia a expressão de genes. Extratos de macroalgas marinhas são o exemplo mais estudado nessa categoria. Ali et al. (2019) mostraram que compostos de Ascophyllum nodosum alteram a transcrição de genes da biossíntese hormonal na planta hospedeira. O hormônio produzido é da planta, não do produto.
A confusão entre as três categorias não é só semântica. Cada uma funciona por mecanismo diferente, em condição diferente e com evidência diferente. Você pode aprofundar como cada um desses compostos se relaciona com a fisiologia vegetal no blog da Agroadvance.
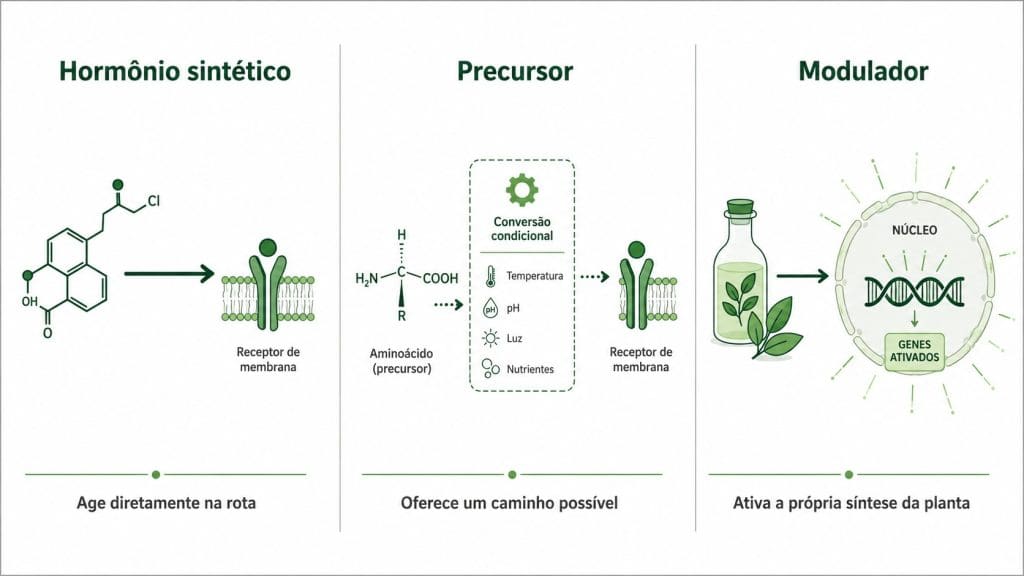
Figura 1. Os três mecanismos que o mercado trata como equivalentes: hormônio sintético age diretamente na rota; precursor oferece um caminho possível; modulador ativa a própria síntese da planta.
Hormônio sintético: ferramenta de precisão, não solução universal
Reguladores vegetais sintéticos têm eficácia documentada e são ferramentas legítimas do manejo agronômico. O ponto não é questionar sua validade. É entender que a janela de uso existe, é previsível e está bem descrita na literatura.
Mendes Fagherazzi et al. (2018) testaram trinexapac-etil em milho e mostraram que o estádio de aplicação determina o resultado. O produto inibe a biossíntese de ácido giberélico ativo na etapa da 3-beta-hidroxilase, reduzindo o alongamento de internódios.
Dentro da janela correta, esse efeito é agronômico e desejável. Para entender como a giberelina atua no crescimento de plantas, o Agrotécnico traz o mecanismo detalhado. Aplicações sequenciais entre V2 e V7, ou tardias após V8, resultaram em redução do comprimento de espiga e da massa de mil grãos. O produto não falhou. Ele foi usado fora da condição em que a planta responde de forma prevista.
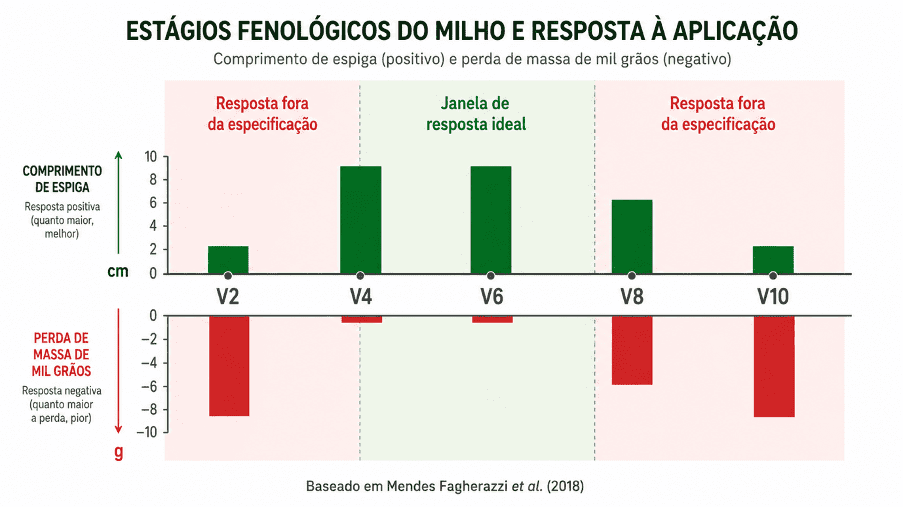
Gráfico 3. O estádio de aplicação do trinexapac-etil determina o resultado agronômico em milho. Mendes Fagherazzi et al. (2018) mostraram que aplicações fora da janela ideal resultam em redução do comprimento de espiga e da massa de mil grãos. O produto não falhou, mas foi aplicado fora da condição em que a planta responde de forma prevista.
Em soja, Silva et al. (2020) avaliaram herbicidas com efeito morfológico e reguladores de crescimento em pós-emergência. Os resultados mostraram que aplicações fora do intervalo V3-V4 geraram resposta diferente da esperada.
O cloreto de mepiquate inibe a formação de ent-caureno de forma irreversível. Se a cultura já enfrenta restrição hídrica no momento da aplicação, o fluxo de carbono para tecidos reprodutivos pode ser comprometido.
Na cana-de-açúcar, Moore e Osgood (1989) documentaram a eficácia do etefom na elevação do Açúcar Total Recuperável ao induzir amadurecimento via etileno. A aplicação com antecedência excessiva à colheita ou em solo com déficit hídrico grave resulta em inibição do crescimento e queda na tonelagem de colmos.
O ganho industrial não compensa a perda de biomassa quando a planta não está em condição de responder adequadamente.
O princípio que une esses três exemplos é o mesmo de Trewavas: a resposta depende da condição da planta naquele momento. Hormônio sintético bem posicionado é uma das ferramentas mais previsíveis disponíveis. Mal posicionado, o efeito vai em outra direção.
Precursor: o que precisa estar resolvido antes
O triptofano é o exemplo central quando se fala em precursor hormonal no mercado de bioestimulantes. A lógica parece direta: triptofano é precursor de auxina, auxina estimula enraizamento e crescimento, logo o produto com triptofano gera resposta de crescimento. Essa cadeia tem dois elos que raramente aparecem na conversa técnica.
O primeiro é o cofator enzimático. Takaki e Kushizaki (1970) documentaram em plântulas de milho com deficiência de zinco o acúmulo de triptofano livre nos tecidos. A rota de conversão para ácido indol-3-acético (IAA) não avançava porque a triptofano sintase, enzima que opera nessa via, requer zinco como cofator estrutural.
O precursor estava presente. O cofator não. O resultado foi encurtamento de internódios e arquitetura foliar reduzida, sintomas que na prática refletem ausência de auxina ativa. Para entender melhor como a auxina age nas respostas das plantas, o Agrotécnico detalha esse mecanismo em profundidade.
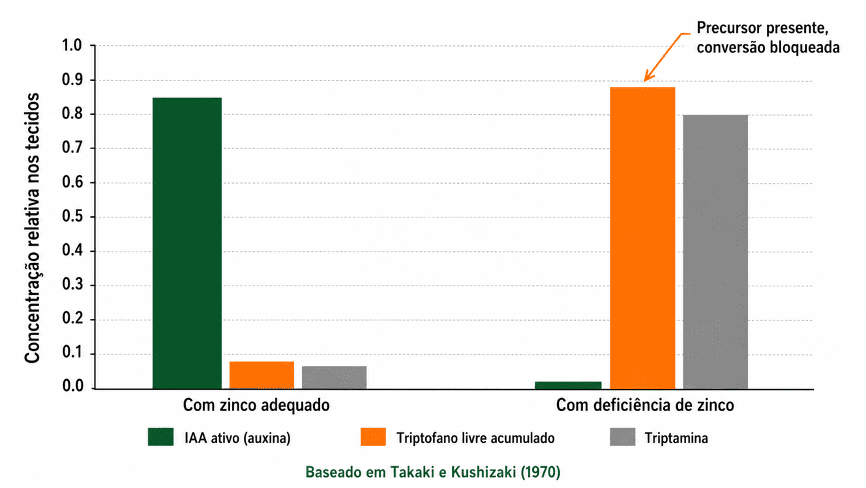
Gráfico 2 - Em soja sob estresse oxidativo, o triptofano exógeno é direcionado para atividade antioxidante, elevando SOD, CAT e APX em vez de seguir para a biossíntese de auxina. Queiroz et al. (2023) demonstraram que a condição fisiológica da planta define o destino metabólico do precursor, não a dose aplicada.
O segundo elo é a demanda da planta naquele momento. Queiroz et al. (2023) avaliaram o efeito do triptofano exógeno em plântulas de Glycine max sob estresse oxidativo. As plantas captaram o aminoácido e o direcionaram para atividade antioxidante, com aumento nas enzimas superóxido dismutase, catalase e ascorbato peroxidase. A proteção ocorreu, mas síntese de auxina não foi o resultado.
Pelagio-Flores et al. (2011) confirmaram o mecanismo: quando a planta está sob estresse, parte do triptofano é desviada para serotonina e melatonina, que atuam na proteção celular contra dano oxidativo.
Isso não invalida o uso de bioestimulantes com triptofano. Significa que, para que o precursor percorra a rota desejada, duas condições precisam estar resolvidas antes da aplicação. O zinco foliar precisa estar na faixa adequada nos tecidos. E a planta precisa estar em condição de crescimento, não de defesa.
Você pode entender melhor como os aminoácidos para plantas atuam em cada rota metabólica no blog da Agroadvance.
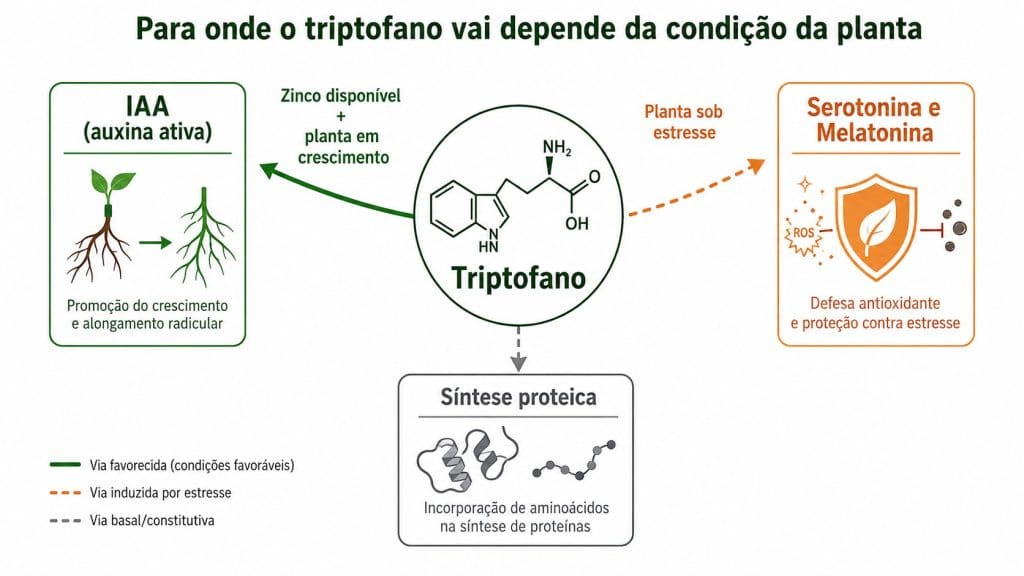
Figura 2. Para onde o triptofano vai depende da condição da planta. Com zinco disponível e planta em crescimento, a rota favorecida é o IAA. Sob estresse, o desvio ocorre para serotonina e melatonina.
Modulador: a planta produz os seus próprios hormônios
Extratos de macroalgas marinhas são a categoria de moduladores com maior volume de evidência publicada nos últimos dez anos. Quando esses produtos são posicionados como hormonais, o argumento ainda sugere, com frequência, que sua eficácia vem de hormônios presentes na matriz algácea. A biologia molecular indica um mecanismo diferente.
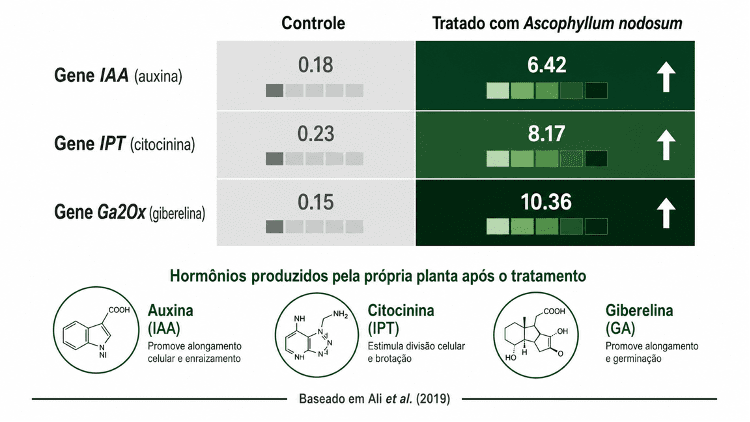
Ali et al. (2019) avaliaram o extrato alcalino de Ascophyllum nodosum em tomate e pimentão em ambiente tropical. A análise de expressão gênica mostrou ativação de genes da biossíntese de auxina endógena, aumento na expressão da isopentenil transferase (IPT), enzima ligada à síntese de citocininas, e incremento da Ga2Ox, reguladora da via de giberelinas. A planta produziu mais dos seus próprios hormônios após o tratamento. Os hormônios do extrato não eram a fonte.
Trivedi et al. (2021) analisaram a expressão gênica foliar de milho tratado com Kappaphycus alvarezii sob déficit hídrico. O extrato ativou mais de 380 genes, incluindo os ligados ao transporte de nitrato e ao maquinário fotossintético. Reduziu a atividade do gene NCED4, que participa da síntese de ácido abscísico sob estresse. As plantas tratadas mantiveram atividade fotossintética em condição que reduziu severamente a eficiência do grupo controle.
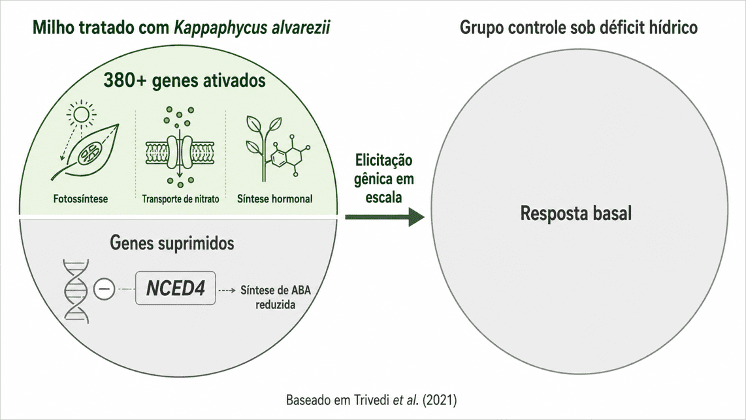
Legenda: O extrato de Kappaphycus alvarezii ativou mais de 380 genes em milho sob déficit hídrico, incluindo os ligados ao maquinário fotossintético e ao transporte de nitrato, e reduziu a atividade do gene NCED4, responsável pela síntese de ABA. Trivedi et al. (2021) demonstraram que o mecanismo de ação dos moduladores algáceos opera por reprogramação da expressão gênica, não por entrega direta de hormônio.
O mecanismo opera por ativação gênica. Polissacarídeos sulfatados, betaínas e florotaninos presentes nos extratos são percebidos por receptores de membrana como sinais moleculares.
Essa percepção induz reprogramação da expressão de genes. O resultado fisiológico é real, documentado e reproduzível. O Agrotécnico traz uma análise detalhada dos tipos de extratos de algas na agricultura e suas diferenças de composição e mecanismo.
Produtos registrados como nutrição foliar podem conter os ativos que modulam essas rotas fisiológicas. O registro descreve a categoria regulatória, não todos os processos que ocorrem na planta após a absorção. A resposta fisiológica acontece pelo mecanismo, independente da denominação no rótulo.
Os metabólitos secundários das plantas envolvidos nesse processo de ativação têm artigo específico no blog da Agroadvance.

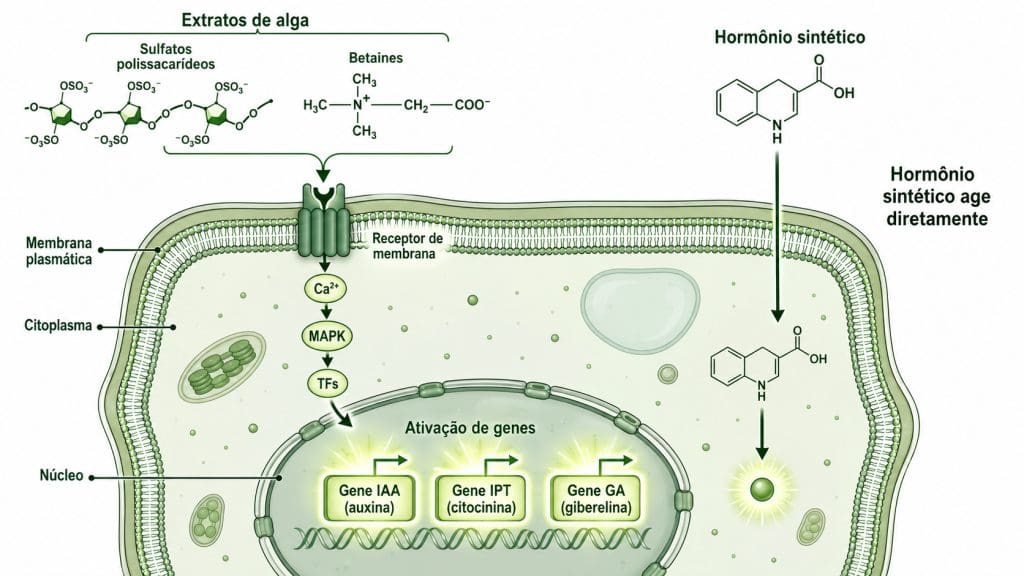
Figura 3. Mecanismo de ação dos extratos de alga por ativação gênica (via Ca²⁺, MAPK e fatores de transcrição) versus ação direta do hormônio sintético na célula vegetal.
Balanço hormonal: ler a planta antes de intervir
Fitohormônios não operam em rotas isoladas. Auxina, citocinina, ácido abscísico (ABA), etileno e giberelinas se regulam mutuamente em ciclos contínuos de ativação e inibição. Qualquer intervenção que altere uma via vai ter seu efeito final moldado pela condição da planta nas outras.
No desenvolvimento radicular, auxina e citocinina precisam coexistir em equilíbrio. A auxina produzida no meristema apical desce via transportadores PIN até as raízes e inicia primórdios de raízes laterais. A citocinina endógena radicular modula a densidade desses transportadores e limita os pulsos de auxina. Excesso de citocinina reduz a ramificação. Déficit de citocinina permite crescimento sem arquitetura funcional para exploração do solo. O ácido abscísico entra nesse balanço como regulador adicional, especialmente sob déficit hídrico, redirecionando o crescimento radicular para camadas mais profundas do perfil de solo.
O exemplo mais direto para a soja está na nodulação. Guinel e Geil (2002) revisaram a relação entre etileno e Bradyrhizobium japonicum e mostraram que o etileno endógeno suprime a infecção bacteriana. Picos de etileno gerados por estresse em V1-V4, por compactação superficial, alagamento temporário ou déficit hídrico nas primeiras semanas após a emergência, reduzem o número de nódulos formados naquela safra. O inoculante foi aplicado corretamente. O problema estava no balanço hormonal gerado pelo estresse precoce, dias antes de qualquer sintoma visível. Como mostram estudos sobre bioestimulantes para déficit hídrico em soja, a janela V1-V4 é crítica para a estabilidade fisiológica que sustenta a nodulação. A fixação biológica de nitrogênio pode suprir entre 50 e 70% da demanda da soja. Comprometer a nodulação nessa janela tem consequência direta no resultado final.
No estádio reprodutivo, o pegamento de vagens envolve o mesmo princípio. O ovário em desenvolvimento produz auxina que flui pelo pedúnculo e mantém a zona de abscisão insensível ao etileno. Quando estresse energético reduz o fluxo de fotoassimilados, a síntese local de auxina cai, a zona de abscisão se torna sensível ao etileno, e a estrutura reprodutiva é descartada. O sintoma visível no campo, flor ou chumbinho no solo, é o estágio final de um processo iniciado dias antes (Schaller et al., 2015).
Reconhecer esse balanço como sistema de leitura da lavoura é o que separa o diagnóstico do protocolo cego. Você encontra mais sobre o papel dos hormônios vegetais nesse equilíbrio no blog da Agroadvance.
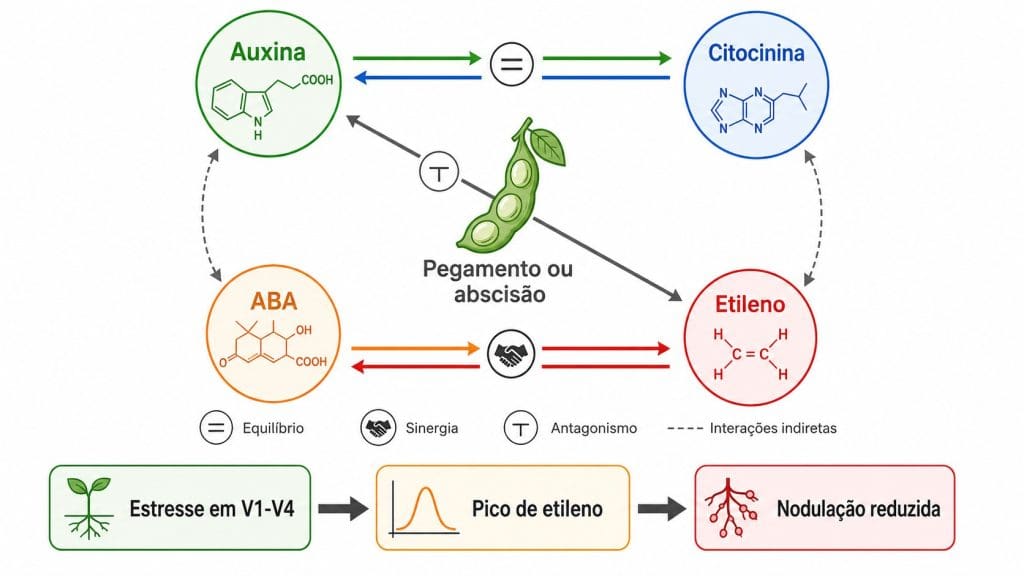
Figura 4. Interações entre auxina, citocinina, ABA e etileno no pegamento ou abscisão de vagens em soja. A linha do tempo inferior mostra a consequência do pico de etileno por estresse precoce em V1-V4 sobre a nodulação.
As três perguntas antes de qualquer aplicação
Com os mecanismos descritos, é possível traduzir esse conhecimento em critério operacional. Três perguntas organizam a avaliação de qualquer produto posicionado como hormonal antes da decisão de uso.
Qual mecanismo está sendo proposto?
É hormônio sintético com ação direta em rota enzimática conhecida? É precursor que depende de conversão mediada por cofator? É modulador que atua por ativação gênica? Cada resposta define a condição de uso. Hormônio sintético exige janela fenológica precisa. Precursor exige diagnóstico nutricional prévio, especialmente zinco. Modulador tem janela mais ampla mas depende de uma condição mínima de equilíbrio fisiológico para ativar a resposta.
Em qual fase da cultura e em qual condição a planta está pronta para responder?
Um produto indicado para enraizamento tem perfil diferente de um indicado para floração ou enchimento de grãos. Engel et al. (2024) testaram hidrolisado proteico em soja via tratamento de sementes e aplicação foliar e registraram alterações no metabolismo de nitrogênio e nos componentes de rendimento com delineamento em blocos casualizados. O resultado foi obtido em condição específica de estádio e dose. Extrapolar para qualquer fase sem essa referência é suposição.
Como a eficácia foi demonstrada?
Bioensaio de elongação radicular, dado de campo com repetição, publicação em periódico revisado por pares? Inca-Torres et al. (2023) descrevem o bioensaio de elongação radicular como método de triagem confiável. Produtos com bioatividade real promovem elongação superior ao controle. Produtos degradados ou de baixa qualidade não produzem esse efeito mesmo com composição analítica idêntica a produtos eficazes. O dado de campo com delineamento adequado continua sendo o critério definitivo.
Quem responde as três perguntas antes de recomendar está num nível de conversa técnica que a maioria do mercado ainda não alcançou. Entender os bioestimulantes e seus modos de ação com esse nível de critério é o que a Agroadvance propõe no seu conteúdo técnico.
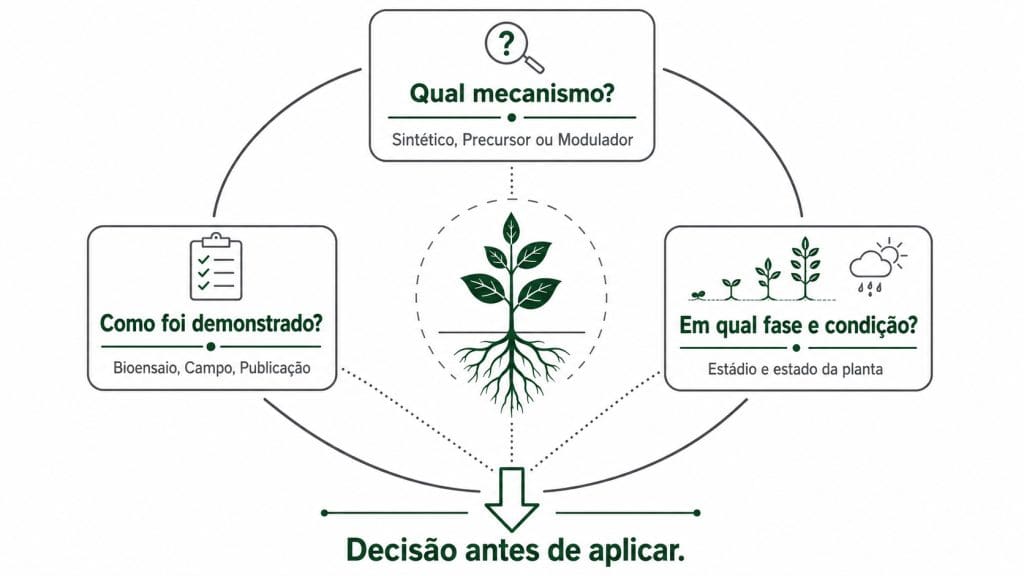
Figura 5. As três perguntas que orientam a decisão de uso de qualquer produto posicionado como hormonal.
Conclusão
A resposta da planta a hormônios, precursores e moduladores não começa no produto. Começa na condição da planta naquele momento. Trewavas (1982) demonstrou que a capacidade de resposta da planta é o fator que determina se o sinal resulta em efeito mensurável. O campo confirma isso safra após safra, em lavouras com o mesmo produto e resultados diferentes.
Hormônio sintético bem posicionado entrega resultado previsível. Precursor com rota desbloqueada contribui para a biossíntese endógena. Modulador com evidência de ativação gênica prepara a planta para produzir os seus próprios sinais hormonais. Os três mecanismos são legítimos. Nenhum funciona fora da condição que o sustenta.
As três perguntas
— qual mecanismo, em qual fase e condição, como foi demonstrado
— são o instrumento que transforma esse conhecimento em decisão de campo.
O técnico que as responde antes de recomendar não espera o resultado para corrigir. Ele lê a planta, identifica a condição e posiciona a intervenção onde ela está pronta para responder.
O produto não precisa ser hormônio para favorecer uma resposta regulada pela planta.
Se você quer aprofundar os processos fisiológicos que sustentam essas decisões em soja, milho, café e cana-de-açúcar, o Guia Técnico de Fisiologia Vegetal da Agroadvance conecta os mecanismos de absorção, transporte e sinalização hormonal com manejo prático de campo.
Referências
ALI, O.; RAMSUBHAG, A.; JAYARAMAN, J. Biostimulatory activities of Ascophyllum nodosum extract in tomato and sweet pepper crops in a tropical environment. PLOS ONE, v. 14, n. 5, e0216710, 2019. DOI: 10.1371/journal.pone.0216710.
BRADFORD, K. J.; TREWAVAS, A. J. Sensitivity thresholds and variable time scales in plant hormone action. Plant Physiology, v. 105, n. 4, p. 1029-1036, 1994. DOI: 10.1104/pp.105.4.1029.
ENGEL, D. C. H. et al. Soybean productivity is increased with protein hydrolysate seed and foliar application. Preprints, 2024. DOI: 10.20944/preprints202406.1387.v1.
GUINEL, F. C.; GEIL, R. D. A model for the development of the rhizobial and arbuscular mycorrhizal symbioses in legumes and its use to understand the roles of ethylene in the establishment of these two symbioses. Canadian Journal of Botany, v. 80, n. 7, p. 695-720, 2002. DOI:10.1139/b02-066
INCA-TORRES, A. R. et al. Enzymatic preparation of mushroom by-product protein hydrolysates. Waste and Biomass Valorization, v. 14, n. 9, p. 3089-3104, 2023. DOI: 10.1007/s12649-023-02157-w.
MENDES FAGHERAZZI, M. et al. Phenological sensitivity of two maize cultivars to trinexapac-ethyl. Planta Daninha, v. 36, n. 1, e017154739, 2018. DOI: 10.1590/s0100-83582018360100012.
MOORE, P. H.; OSGOOD, R. V. Prevention of flowering and increasing sugar yield of sugarcane by application of Ethephon (2-Chloroethylphosphonic acid). Journal of Plant Growth Regulation, v. 8, n. 3, p. 205-210, 1989. Disponivel em : Journal growth regulation
PELAGIO-FLORES, R. et al. Serotonin, a tryptophan-derived signal conserved in plants and animals, regulates root system architecture probably acting as a natural auxin inhibitor in Arabidopsis thaliana. Plant and Cell Physiology, v. 52, n. 3, p. 490-508, 2011. DOI: 10.1093/pcp/pcr006
QUEIROZ, R. B. et al. Effect of exogenous tryptophan on primary metabolism and oxidative stress and their relationship with seedling germination and vigor of Glycine max L. Agronomy, v. 13, n. 6, p. 1609, 2023. DOI: 10.3390/agronomy13061609.
SCHALLER, G. E. et al. Cross-talk between environmental stresses and plant metabolism during reproductive organ abscission. Journal of Experimental Botany, v. 66, n. 7, p. 1707-1719, 2015. DOI: 10.1093/jxb/eru533.
SILVA, A. F. M. et al. Post-emergence application of herbicides and growth regulators on soybean growth and agronomic performance. Journal of Crop Science and Biotechnology, v. 23, p. 253-258, 2020. DOI: 10.1007/s12892-020-00033-w.
TAKAKI, H.; KUSHIZAKI, M. Accumulation of free tryptophan and tryptamine in zinc deficient maize seedlings. Plant and Cell Physiology, v. 11, n. 5, p. 793-804, 1970. DOI: 10.1093/oxfordjournals.pcp.a074567.
TREWAVAS, A. How do plant growth substances work? Plant, Cell & Environment, v. 4, n. 3, p. 203-228, 1981. DOI: 10.1111/j.1365-3040.1991.tb01366.xDigital Object Identifier (DOI)
TREWAVAS, A. Growth substance sensitivity: the limiting factor in plant development. Physiologia Plantarum, v. 55, n. 1, p. 60-72, 1982. DOI: 10.1111/j.1399-3054.1982.tb00285.x.
TRIVEDI, K. et al. Transcriptional analysis of maize leaf tissue treated with seaweed extract under drought stress. Frontiers in Sustainable Food Systems, v. 5, p. 774978, 2021. DOI: 10.3389/fsufs.2021.774978.
Como citar este artigo
BUENO, D. R. Hormônios vegetais: sintético, precursor e modulador no campo. Agrotécnico Publicado: 07 de Junho de 2026. Disponível em: https://agrotecnico.com.br/hormonios-vegetais-sintetico-precursor-e-modulador-no-campo/. Acesso: dd Mmm. aaaa.
Sobre o autor

Deyvid Rodrigues Bueno
Fundador do Agrotécnico.com.br
Gerente de Produtos · ICL América do Sul · Gestão de Produtos (Business Behavior Institute) · MBA em Gestão, Empreendedorismo e Marketing (PUCRS) · Especialista em Solos e Nutrição de Plantas (FAZU) · Engenheiro Agrônomo (UFMT) · Técnico Agrícola (IFRO)




0 Comentários