

O estresse pode ser definido como qualquer condição ambiental que impeça as plantas de atingir todo o seu potencial genético. Por exemplo, uma diminuição na intensidade da luz levará a uma diminuição na atividade fotossintética e, ao mesmo tempo, o fornecimento de energia das plantas também diminuirá. Nessas condições, as plantas podem compensar de duas maneiras: desacelerando a biossíntese, reduzindo assim sua taxa de crescimento, ou usando reservas de alimentos armazenadas na forma de amido (Taiz et al., 2017).
Da mesma forma, a redução na quantidade de água disponível também pode ter um efeito prejudicial no crescimento. Uma forma de compensar a queda no potencial hídrico é fechar os estômatos, reduzindo assim a perda de água causada pela transpiração. No entanto, o fechamento dos estômatos também reduz a absorção de dióxido de carbono pelas folhas, reduzindo a fotossíntese e inibindo o crescimento.
As plantas estão constantemente expostas a diferentes fatores de estresse abiótico sem qualquer proteção. Por outro lado, no manejo dos animais, mesmo que intuitivamente, eles podem escapar das condições de estresse abiótico direto.
Está claro que, devido a essa natureza, as plantas desenvolveram mecanismos moleculares únicos para lidar com diferentes fatores de estresse. Mesmo assim ainda existem diferenças nos mecanismos de tolerância ao estresse das plantas com características morfológicas adaptativas em certas plantas que permitem que elas evitem estressores. A única opção para as plantas é mudar sua fisiologia, mecanismos metabólicos, expressão gênica e atividades de desenvolvimento para lidar com os efeitos do estresse.
Dessa forma vemos que as plantas possuem mecanismos únicos e complexos para tolerar o estresse abiótico. Plantas com maior tolerância, resistência, proteção e mecanismos de adaptação podem sobreviver, mas outras plantas não.

As tensões abióticas são comumente causadas pela seca, salinidade, alta ou baixa temperatura, deficiência ou excesso de nutrientes, metais pesados, poluentes, etc., tanto indiretamente como em combinações.
O estresse causado por fatores abióticos altera o metabolismo das plantas levando a efeitos negativos no crescimento, desenvolvimento e produtividade das plantas e ao se prolongar por mais tempo, pode levar a uma carga metabólica insuportável sobre as células, levando a uma redução do crescimento e, em casos extremos, resulta na morte das plantas.
Interferencias do Estresse no desenvolvimento
Sabemos que no decorrer do percurso natural do crescimento das plantas, elas experimentam algum grau de estresse. Para combater esses estresses, as plantas apresentam vários mecanismos que as fazem suportar o estresse com a formação de novas moléculas e mecanismos moleculares de tolerância ao estresse.
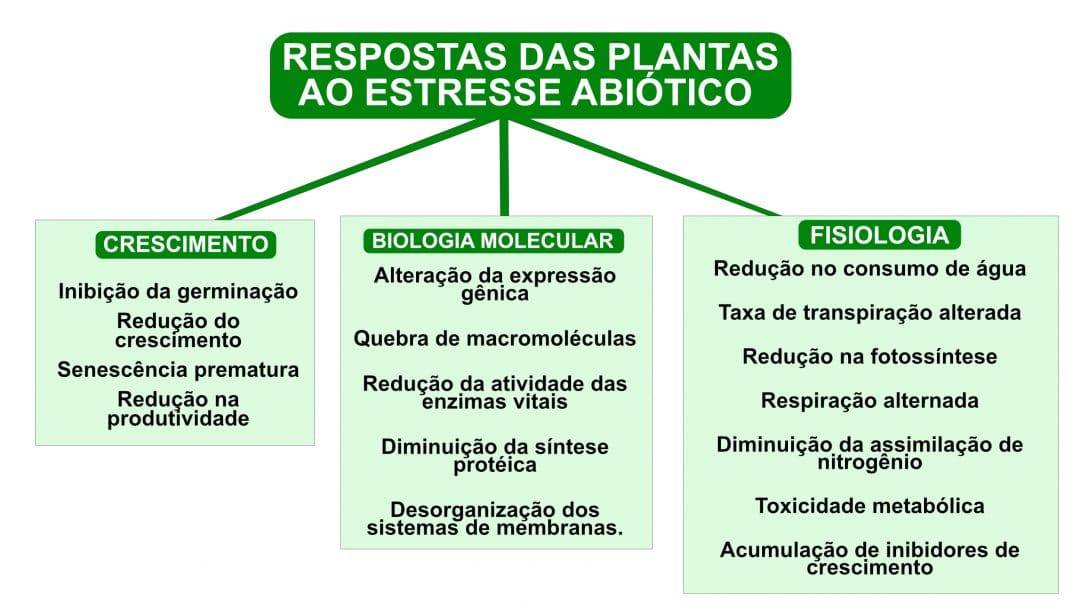
Em cada alteração por resposta ao estresse, há uma série de mecanismos de tolerância que são manifestados para diminuir o seu dano potencial:
- Ativação de fatores de sinalização
- Alteração da expressão gênica
- Acumulação de solutos compatíveis
- Síntese de proteínas de estresse
- Metabolismo antioxidativo aprimorado
- Homeostase de íons e compartimentação
- Transporte de membrana facilitado
- Acumulação de poliaminas
- Ajuste do equilíbrio hormonal
Espécies Reativas de Oxigênio
A resposta para qualquer tipo de estresse pode variar em sua generalidade de ações porém sempre que a planta percebe qualquer efeito estressor uma resposta metabólica padrão ocorre : Formação de Espécies Reativas de Oxigênio, ou EROs.

Para que que haja um dano significativo ao organismo vegetal é necessário que ocorra de um desequilíbrio entre a geração de compostos oxidantes (EROs) e a atuação dos sistemas de defesa antioxidante. A geração de radicais livres e/ou espécies reativas não radicais é resultante do metabolismo de oxigênio.
O sistema de defesa antioxidante tem a função de inibir e/ou reduzir os danos causados pela ação deletéria dos radicais livres e/ou espécies reativas não radicais. Esse sistema, usualmente, é dividido em enzimático (superóxido dismutase, catalase e glutationa peroxidase) e não-enzimático. No último caso, é constituído por grande variedade de substâncias antioxidantes.

Estresse vegetal – EROs e Sistema antioxidante , Linkedin Agrotécnico
Estresse hídrico
O estresse hídrico nas plantas é um grande problema para reduzir a produtividade agrícola, especialmente nas regiões tropicais, semiáridas e áridas do mundo.
Ao nível celular esses déficits de água geram a concentração de solutos, perda de turgor, alteração no volume celular, interrupção de gradientes potenciais de água, alteração na integridade da membrana, desnaturação de proteínas e vários componentes fisiológicos e moleculares.
Os efeitos do estresse dependem do grau e da duração do estresse, do estágio de desenvolvimento da planta, da capacidade genotípica das espécies e das interações ambientais.
A nível molecular a planta provoca a indução gênica levando à formação de produtos compostos como a prolina, glicina betaina e outros, que podem atuar para manter a função celular através da proteção das estruturas celulares e ajustes osmóticos.

A concentração de ácido abscísico aumenta sob estresse hídrico, bem como sob algumas outras tensões abióticas. De fato, o ácido abscísico é considerado como um "hormônio do estresse", embora possa servir a várias outras funções na ausência de estresse.
A compreensão das funções dos vários produtos genéticos formados, que normalmente estão envolvidos no ajuste osmótico, proteção e reparo das estruturas celulares, é de grande valor na avaliação dos mecanismos de tolerância ao estresse hídrico e para desenvolver plantas tolerantes ao estresse hídrico.
A Resposta das plantas ao frio
A manutenção de temperaturas foliares favoráveis é crucial para o crescimento vegetal, porque a fotossíntese máxima ocorre dentro de uma faixa de temperatura relativa mente estreita. O pico da taxa fotossintética em uma faixa de temperaturas é o ideal térmico fotossintético. Quando a temperatura ótima para determinada planta é ultrapassada, as taxas fotossintéticas decrescem novamente. O ideal térmico fotossintético reflete componentes bioquímicos, genéticos (adaptação) e ambientais (aclimatação).
A aclimatação a frio e a sobrevivência no inverno em plantas está fortemente associada a reação da fotossíntese a baixa temperatura e ao armazenamento de carboidratos solúveis, como sacarose e rafinose. Durante o estresse por congelamento, a formação de cristal de gelo extracelular provoca desidratação celular.
As plantas sujeitas a temperaturas de congelamento devem enfrentar a formação de cristais de gelo, tanto no âmbito extracelular quanto no intracelular. A formação de cristais de gelo intracelular quase sempre se mostra letal à célula. No entanto, a água no apoplasto é relativamente diluída e, portanto, tem um ponto de congelamento mais alto do que a do simplasto mais concentrado. Como consequência, cristais de gelo tendem a se formar no apoplasto e em traqueídes e vasos, ao longo dos quais o gelo pode se propagar rapidamente. A formação de cristais de gelo diminui o potencial hídrico do apoplasto, que se torna mais negativo que o do simplasto. Água não conge- lada dentro da célula se move para baixo nesse gradiente, em direção aos cristais de gelo nos espaços intercelulares. À medida que a água deixa a célula, a membrana plasmática contrai-se e afasta-se da parede celular. Durante esse processo, a membrana plasmática, enrijecida pela temperatura baixa, pode ficar danificada. Quanto mais baixa a temperatura, mais água se desloca para baixo nesse gradiente em direção à água congelada. Por exemplo, a –10 graus celsos, o simplasto perde cerca de 90% de sua água osmoticamente ativa para o apoplasto. Nesse sentido, o estresse pelo congelamento tem muito mais em comum com o estresse pela seca. Como ocorre com o estresse pela seca, as células que já estão desidratadas, como nas sementes e nos grãos de pólen, têm menos probabilidade de passarem por outra desidratação pela formação de cristais extracelulares.
Adaptação e resitência ao estresse pelo frio
Plantas de tabaco transgênicas que acumulam altos níveis de prolina, frutano ou glicina betaína exibem tolerância ao frio (Konstantinova et al., 2002; Parvanova et al., 2004). A superexpressão de glutationa S-transferase / glutationa peroxidase faz com que as plantas tolerem o estresse pelo frio (Roxas et al., 1997).
Muitas variedades de trigo e colza mostram uma relação paralela entre o teor de açúcar e a tolerância ao congelamento.
Os principais açúcares que melhoram a resistência ao frio são sacarose, glicose, frutose, sorbitol, manitol e rafinose. Presume-se que o açúcar substitua a água e reduz o grau de desidratação causado pelo congelamento.
Prolina, arginina, treonina e lisina exógenas protegem a membrana tilacóide da inativação por congelamento. A prolina e a glicina betaína são consideradas crioprotetoras.
As plantas utilizam múltiplos caminhos de sinalização e sinais para mediar suas respostas de aclimatação. Apesar de vários apontamentos na literatura para as vias de sinalização para a resposta a esse tipo de estresse é a combinação específica de vários componentes da rede de sinalização acoplados a fatores espaciais e de temporal que, em última instância, resultam em um aumento da resistência ao inverno.
Há uma grande confusão quanto ao papel da ABA no desenvolvimento e manutenção da tolerância ao congelamento, o que levou à sugestão de que existem dois caminhos; um caminho independente da ABA e um caminho dependente da ABA. Esta evidência é baseada na constatação de que o ABA em temperaturas não aclimatadas não consegue regular vários genes associados ao frio.
Muitos aspectos do desenvolvimento de plantas, incluindo a tolerância ao estresse, são regulados por interações antagônicas entre hormônios vegetais, por exemplo:
Ácido abscísico (ABA) e giberelinas (Gomez-Cadenas et al., 2001);
Auxinas e citoquininas (Coenen et al., 2003);
E por interações sinérgicas, por exemplo, ABA e ácido jasmônico (JA) (Wilen et al., 1994).
O ácido abscísico, um inibidor de crescimento, induz a aclimatação a frio em uma ampla gama de plantas, enquanto as giberelinas são promotoras de crescimento e neutralizam o efeito do ABA. Análises recentes de mutantes Arabidopsis demonstraram fortes interações entre ABA e outros caminhos de sinalização - incluindo auxina, açúcar e etileno (Lu e Fedoroff, 2000, Gazzarrini e McCourt, 2003).
Assim, os hormônios compreendem uma rede muito complexa de moléculas de sinalização a nível celular. Isto levou à sugestão de que as respostas de fito hormônios não podem ser reduzidas a simples caminhos lineares que conectam entradas e saídas, mas são mais provavelmente por redes interativas.
Conclusão
Estresses abióticos, de maneira geral despertam respostas muito parecidas no metabolismo vegetal. Isso quando vemos a produção de EROs e demais ajustes hormonais. Para responder aos vários tipos de estresse abióticos as plantas produziram ao longo do tempo várias estratégias adaptativas. Neste texto vimos algumas delas.




0 Comentários
Trackbacks/Pingbacks